Ndërtesa e re e kishës protestante në Austri
Në Moskë, si në të gjithë Rusinë, jetojnë njerëz të shumë feve. Mes banorëve të kryeqytetit ka edhe protestantë. Ata nuk janë...
Grupet C dhe G. Proteina G është e ngjashme me proteinën A, por ndryshon në specifikë. Proteina G ka një peshë molekulare prej 58 kDa (në rastin e proteinës C40) ose 65 kDa (në rastin e proteinës G148). Proteina G lidhet me rajonin α të antitrupave dhe për këtë arsye përdoret gjerësisht për pastrimin e imunoglobulinave. Molekulat e proteinës G gjithashtu lidhin albuminën.
Ka proteina të tjera bakteriale që lidhin imunoglobulinat - proteina A, proteina A/G dhe proteina L. Këto proteina përdoren për pastrimin, imobilizimin dhe izolimin e imunoglobulinave. Këto proteina lidhëse të antitrupave kanë profile të ndryshme të lidhjes së imunoglobulinave.
Fondacioni Wikimedia.
Shihni se çfarë është "Proteina G" në fjalorë të tjerë:
- (eng. proteina A) është një proteinë me peshë molekulare 40–60 kDa, e izoluar nga sipërfaqja e murit qelizor të Staphylococcus aureus (Staphylococcus aureus). Proteina A përdoret në kërkimet biokimike, pasi lidh mirë shumë... ... Wikipedia Gawk at Arab squirrels.. Fjalor sinonimesh ruse dhe shprehje të ngjashme. nën. ed. N. Abramova, M.: Fjalorët Ruse, 1999. Proteina globulin, histon, proteinoid, protein, trup proteinik, protamine, proteid Fjalor i sinonimeve ruse ...
Fjalor sinonimish
Fjalori shpjegues i Ushakovit Fjalor sinonimish
Fjalori shpjegues i Ushakovit Fjalor sinonimish
1. PROTEIN1, ketri, mashkull. (biol. kim.). Përbërësi më i rëndësishëm i trupit të kafshëve dhe bimëve; njëjtë si albumina dhe proteina. 2. PROTEIN2, ketri, mashkull. 1. Predha konveks opake e syrit. || vetëm shumës Sytë (të thjeshtë). Nxirrni të bardhët. Proteinat...... proteina C - Proteina, serine proteaza, e sintetizuar nga qelizat e mëlçisë; antikoagulant, është frenues i faktorëve të koagulimit të gjakut Va dhe VIIIa; frekuenca e heterozigoteve për mungesë B.C në popullatat njerëzore arrin në 1/200 kjo anomali shoqërohet me rritje të... ...
PROTEINA, KOMPONIM organik që përmban shumë AMINOACIDE të lidhura me lidhje peptide kovalente. Molekulat e proteinave përbëhen nga zinxhirë polipeptidikë. Ka rreth 20 aminoacide të ndryshme në QELIZAT e gjalla. Për faktin se në çdo... ... Fjalor enciklopedik shkencor dhe teknik
PROTEINA, lka, burri. Një substancë organike me molekulare të lartë që siguron funksionet vitale të organizmave të kafshëve dhe bimëve. | adj. proteina, oh, oh. Ushqimi i proteinave (përmbajtja e lartë e proteinave). II. PROTEINA, lka, burri. 1. Pjesa transparente e vezës... Fjalori shpjegues i Ozhegovit
Proteina lidhëse e acideve yndyrore (FABP) është një familje e transportuesve të acideve yndyrore dhe substancave të tjera lipofile si eikosanoidet dhe retinoidet. Besohet se këto... ... Wikipedia
1. PROTEINA, lka; m 1. Lëngu transparent që rrethon të verdhën e vezës së një zogu. / Rreth kësaj pjese të një veze pule si ushqim. Pini të papërpunuara b. Të bardhat e rrahura. ◁ Proteina, oh, oh. B. krem (nga e bardha e vezëve). 2. PROTEINA shih 1. Proteinat. 3. PROTEINA shih 2. Proteinat. 4.…… Fjalor Enciklopedik
(anglisht) Proteinat lidhëse të nukleotideve të guaninës, Proteinat lidhëse të nukleotideve guanil janë një familje proteinash të përfshira në sinjalizimin e qelizave eukariote. Proteinat G luajnë rolin e një lloj ndërprerësi: ato mund të kalojnë nga një gjendje joaktive në atë aktive dhe anasjelltas, përkatësisht, duke ndezur ose fikur transmetimin e një sinjali të caktuar brenda qelizës. Këto proteina morën emrin e tyre për aftësinë e tyre për të lidhur nukleotidet guanil. G-anine nukleotidi): në një kompleks me guanozine difosfat (GDP) ato janë joaktive, dhe në një kompleks me guanozinetrifosfat (GTP) janë aktive.
Termi "proteina G" përdoret më shpesh për t'iu referuar proteinave heterotrimerike (të mëdha) që lidhen me GTP të përbëra nga tre nënnjësi α, β dhe γ; Ekziston një klasë tjetër e proteinave të lidhura me GTP - monomere, të cilat nganjëherë quhen proteina të vogla G (supershtëpia e Ras GTPases të vogla), ato janë homologe me nën-njësitë α të atyre të mëdha.
Proteinat heterotrimerike G janë të përfshira në transmetimin e sinjaleve nga receptorët e lidhur me proteinën G. Receptorët e bashkuar me proteinën G (GPCR)- klasa më e madhe e receptorëve qelizorë (për shembull, në Caenorhabditis elegans gjenet e tyre zënë 5% të të gjithë gjenomit). Tek vertebrorët, ata janë përgjegjës për perceptimin e qelizave të një numri hormonesh dhe molekulash të tjera sinjalizuese, si dhe për shqisat kimike (erë dhe shije) dhe fotoreceptimin (vizionin). Është domethënëse që përafërsisht gjysma e barnave të njohura farmaceutike veprojnë përmes receptorëve të lidhur me proteinën G: këto përfshijnë medikamente të njohura, si antihistaminet Claritin (loratadine) dhe antidepresivët Prozac (Fluoxetine), si dhe substanca psikotrope, në veçanti. heroinë, kokainë dhe tetrahidrokanabinol (përbërës aktiv në marihuanë).
Proteinat heterotrimerike G u zbuluan nga Alfred Gilman dhe Martin Rodbell, për të cilat ata morën çmimin Nobel në Fiziologji ose Mjekësi në 1994.
Proteinat hetrotrimerike G përbëhen nga tre nënnjësi: α, β dhe γ. Nën-njësia α përmban një domen të lidhjes dhe hidrolizës GTP që është identik në të gjithë superatdheun e GTPase. Nën-njësia β përmban 7 struktura β të organizuara si tehe të helikës. Nën-njësia γ ndërvepron ngushtë me β-nënnjësinë së bashku ato formojnë një strukturë të vetme funksionale, e cila mund të shkëputet vetëm në rastin e hidrolizës së proteinave; E gjithë proteina G është e ankoruar në membranë nga dy lipide, njëri prej të cilëve është i lidhur në mënyrë kovalente me fundin N të nën-njësisë α, tjetri në fundin C të nën-njësisë γ.
Receptorët e lidhur me proteinën G Receptorët e bashkuar me proteinën G, GPCR)- familja më e madhe e receptorëve qelizorë në eukariotët, duke siguruar perceptimin e hormoneve, neurotransmetuesve, rregullatorëve lokalë, si dhe siguron shikimin, nuhatjen dhe ndjenjën e shijes tek vertebrorët. Rreth 700 gjene GPCR gjenden në gjenomin e njeriut, dhe tek miu, më shumë se 1000 prej këtyre receptorëve i përgjigjen vetëm shqisës së nuhatjes.
Molekulat e sinjalit që veprojnë si ligandë për receptorët e lidhur me proteinën G mund të jenë shumë të ndryshme në natyrën kimike: proteina, peptide të vogla, lipide, derivate të aminoacideve dhe të ngjashme. Përveç kësaj, disa anëtarë të kësaj klase të receptorëve, në veçanti rodopsina, mund të perceptojnë fotonet e dritës. Ndonjëherë ekzistojnë disa GPCR të ndryshme për një molekulë të vetme sinjalizuese, shprehen në lloje të ndryshme qelizash dhe nxisin rrugë të ndryshme sinjalizimi. Për shembull, në trupin e njeriut ka të paktën 9 receptorë të ndryshëm për adrenalinën dhe të paktën 14 për neurotransmetuesin serotonin.
Të gjithë receptorët e lidhur me proteinën G kanë një strukturë të ngjashme: ata përbëhen nga një zinxhir i vetëm polipeptid që përshkon shtresën e dyfishtë lipidike 7 herë. Çdo domen transmembranor përfaqësohet nga një α-helix, i cili përfshin 20-30 aminoacide jopolare. Këto fusha janë të ndërlidhura nga sythe të madhësive të ndryshme të vendosura në të dy anët e membranës plazmatike. GPCR-të janë kryesisht glikoproteina, mbetjet e karbohidrateve të të cilave ndodhen në anën e glikoproteinës. Domenet ndërqelizore të këtyre receptorëve përmbajnë vende ndërveprimi me proteinat G.
Proteinat G shërbejnë për rolin e bashkimit të receptorëve qelizorë me disa molekula efektore, të tilla si enzimat ose kanalet jonike, ndërsa ato veprojnë si ndërprerës molekularë. Në gjendjen joaktive, proteinat G përmbajnë GDP të lidhur me nënnjësinë α.
Transmetimi i sinjalit fillon kur ligandi i duhur vepron në receptorin qelizor, si rezultat i të cilit receptori aktivizohet dhe ndryshon konformacionin. Receptori i aktivizuar ndikon në proteinën G (e cila ose kompleksohet përgjithmonë me të ose lidhet pas aktivizimit), duke bërë që struktura e nën-njësisë α të ndryshojë në atë mënyrë që liron molekulën e GDP të lidhur. Vendin e kësaj molekule e zë shpejt GTP, e cila çon në aktivizimin e proteinës G dhe ndryshimet në strukturën e saj: nën-njësia α humbet afinitetin për kompleksin βγ dhe ajo shpërbëhet. Në një gjendje të tillë të aktivizuar, si nën-njësia α e lidhur me GTP dhe kompleksi βγ mund të kryejnë transmetimin e sinjalit: aktivizojnë disa enzima ose ndikojnë në gjendjen e kanaleve jonike. Nën-njësia α është një GTPazë dhe sapo hidrolizon GTP-në e lidhur me GDP, ajo çaktivizohet menjëherë dhe struktura e prerësit të proteinës G restaurohet. Në këtë mënyrë tingulli fiket. Proteina G e inaktivizuar mund të ndërveprojë me një molekulë të receptorit pasues dhe të ndizet përsëri.
Efikasiteti i transmetimit të një sinjali të veçantë përmes proteinës G varet nga raporti midis përqendrimit të formave aktive, të lidhura me GTP dhe joaktive, të lidhura me GDP. Dhe ky raport, nga ana tjetër, varet nga dy gjendje: konstanta e disociimit të PBB-së dhe konstanta e shkallës së hidrolizës GTP:
Ku
Kjo lidhje konfirmohet nga një tepricë e GTP në medium, si dhe nga lidhja e tij e shpejtë, pothuajse e menjëhershme, me një molekulë "të zbrazët" të proteinës G (d.m.th., jo e lidhur me një nukleotid guanil). Në këtë rast, efikasiteti i transmetimit të sinjalit mund të rregullohet në një nga mënyrat e mëposhtme:
Proteinat G marrin të dhëna nga receptorët e tyre të lidhur, pas së cilës ato aktivizojnë një nga rrugët e sinjalizimit të qelizës.
AMP ciklike (cAMP) është një lajmëtar i dytë i zakonshëm që kontrollon shumë procese në qelizat eukariote. cAMP sintetizohet me ATP nga enzima e madhe transmembranore adenilate ciklaza dhe degradohet nga cAMP fosfodiesteraza. Shumë molekula sinjalizuese ndikojnë në qelizë duke rritur ose ulur përqendrimin e cAMP përmes aktivizimit ose frenimit të adenilate ciklazës. cAMP kryen funksionin e tij si një lajmëtar i dytë duke aktivizuar protein kinazën e varur nga cAMP (protein kinaza A, PKA), e cila nga ana e saj fosforilon shumë proteina në qelizë në mbetjet e serinës dhe treoninës, duke i aktivizuar ose çaktivizuar ato.
Ekzistojnë dy lloje të proteinave G që ndikojnë në ciklazat aktive adenilate: G s (eng. Stimuluese)- stimulimin, aktivizimin e tij dhe rritjen e përqendrimit të cAMP dhe G i (eng. Frenues)- frenues, duke shtypur adenilate ciklazën, por vepron edhe me veprim të drejtpërdrejtë në kanalet jonike. Shembuj të reaksioneve të shkaktuara nga një rritje e varur nga Gs në përqendrimin e cAMP janë:
G-proteinat, të cilat ndikojnë në sinjalizimin qelizor të varur nga cAMP, janë objektiva të toksinave bakteriale:
Këto dy toksina përdoren në studimet biologjike për të përcaktuar se cila përgjigje qelizore ndërmjetësohet nga proteina G s - ose G i.
Shumë receptorë të lidhur me proteinën G veprojnë duke aktivizuar fosfolipazën C-β (PLC-β). Kjo enzimë vepron në fosfolipidin e inozitolit: fosfatidilinositol 4,5 bisfosfat (PI(4,5)P2 ose PIF 2), i pranishëm në sasi të vogla në fletën e brendshme të shtresës së dyfishtë lipidike të membranës plazmatike. Receptorët që aktivizojnë këtë rrugë sinjalizuese zakonisht lidhen me bllokun Gq, i cili aktivizon fosfolipazën C në mënyrë të ngjashme me Gs-bllok-adenilate ciklazën. Fosfolipaza e aktivizuar zbërthen fosfatidilinositol 4,5 bisfosfat në inositol 1,4,5 trifosfat (IP 3) dhe diaciglicerol (DAG). Në këtë fazë, rruga e sinjalizimit degëzon:
Shembuj të reaksioneve qelizore të aktivizimit të fosfolipazës C-β të varur nga proteina G janë:
Shumë proteina G veprojnë duke hapur ose mbyllur kanalet jonike, duke ndryshuar kështu vetitë elektrike të membranës plazmatike.
Për shembull, një rënie në frekuencën dhe forcën e kontraktimeve të zemrës nën ndikimin e acetilkolinës ndodh për shkak të faktit se receptorët muskarinikë të acetilkolinës, pas aktivizimit, ndërveprojnë me G i-bill, nën-njësia e së cilës shtyp aktivitetin e adenilatit. ciklaza, ndërsa kompleksi βγ hap kanalet e kaliumit në membranën plazmatike të muskujve të qelizave kardiake, për shkak të të cilave ulet ngacmueshmëria e tyre.
Proteinat e tjera G rregullojnë aktivitetin e kanaleve jonike në mënyrë indirekte: për shembull, receptorët e shikimit dhe të nuhatjes veprojnë përmes proteinave G, të cilat ndikojnë në sintezën e nukleotideve ciklike, duke mbyllur ose hapur kanalet jonike (kanalet jonike të kontrolluara nga nukleotidet ciklike). Për shembull, të gjithë receptorët e nuhatjes janë të lidhur me proteinën G olf, e cila aktivizon adenilate ciklazën; CAMP që sintetizohet hap kanalet e natriumit, gjë që çon në depolarizimin e membranës dhe gjenerimin e një impulsi nervor (potenciali i receptorit) që transmetohet te neuronet.
Në shufrat e retinës njerëzore, molekula e ndjeshme ndaj dritës është rodopsina. Membrana plazmatike e këtyre qelizave përmban një numër të madh kanalesh kationesh të mbyllura nga cGMP. Në mungesë të stimulimit të dritës, citoplazma e shufrave përmban një sasi të lartë të cGMP, e cila i mban kanalet e kationeve të hapura. Si rezultat, membrana depolarizohet periodikisht dhe ndodh transmetimi sinaptik i impulseve te neuronet. Pas aktivizimit nga drita, rodopsina ndryshon konformimin dhe ndërvepron me transducinën e proteinës G (Gt). Pas kësaj, nën-njësia e saj α aktivizon fosfodiesterazën cGMP, e cila zbërthen cGMP, si rezultat i së cilës kanalet e kationit mbyllen dhe transmetimi sinaptik ndalon. Është ulja e frekuencës së impulseve që vijnë nga qelizat e ndjeshme ndaj dritës, e cila perceptohet nga truri si një ndjesi drite.
Të gjitha proteinat heterotrimerike G ndahen në katër familje kryesore bazuar në sekuencën e aminoacideve të nën-njësisë α:
| Familjet kryesore të proteinave G heterotrimerike bazuar në sekuencën e aminoacideve të nën-njësisë α | |||
|---|---|---|---|
| Familja | Disa anëtarë | Nën-njësia përgjegjëse për efektin | Disa veçori |
| I | G s | α | Aktivizimi i adenilate ciklazës, hapja e kanaleve të kalciumit |
| G olf | α | Aktivizimi i adenilate ciklazës në neuronet e nuhatjes | |
| II | G i | α | Frenimi i adenilate ciklazës |
| βγ | Hapja e kanaleve të kaliumit | ||
| G o | βγ | Hapja e kanaleve të kaliumit, mbyllja e kanaleve të kalciumit | |
| α dhe βγ | Aktivizimi i fosfolipazës C-β | ||
| G t (transducin) | α | Aktivizimi i fosfodiesterazës cGMP në fotoreceptorët e vertebrorëve | |
| III | Gq | α | Aktivizimi i fosfolipazës C-β |
| IV | G 12/13 | α | Aktivizimi i GTP-azave monomerike të familjes Rho që rregullojnë citoskeletin e aktinës |
(Receptorët e lidhur me proteinën C, GPCRs), transmetojnë një sinjal nga lajmëtarët parësorë në objektivat ndërqelizorë duke përdorur kaskadën e proteinës GPCR-^-G-protein^-efektor. Sinjalet kryesore për këta receptorë janë një shumëllojshmëri e gjerë molekulash, për shembull, hormonet me peshë të ulët molekulare dhe neurotransmetuesit (si adrenalina, norepinefrina, acetilkolina, serotonina, histamina), opioidet, peptidet dhe hormonet proteinike (adrenokortikotropina, somaostatina, angiopresinina , gonadotropina, faktori i rritjes epidermale), disa neuropeptide.
E njëjta seri përfshin shumë sinjale kimike të perceptuara nga qelizat shqisore të nuhatjes dhe shijes, dhe dritën, receptori për të cilin është pigmenti i qelizave vizuale ose fotoreceptore rodopsina.
Duhet të kihet parasysh se i njëjti sinjal primar mund të inicojë transmetimin e sinjalit përmes disa (ndonjëherë më shumë se 10) GPCR të ndryshme, kështu që nëse numri i sinjaleve të jashtme për GPCR është disa dhjetëra, atëherë dihen më shumë se 200 receptorë të tillë.
Me gjithë diversitetin e tyre, GPCR-të janë proteina integrale monomere të membranës, zinxhiri polipeptid i të cilave kalon shtatë herë në membranën qelizore. Në të gjitha rastet, vendi i receptorit përgjegjës për ndërveprimin me sinjalin parësor është i lokalizuar në anën e jashtme të membranës, dhe rajoni në kontakt me proteinën G është në anën e saj citoplazmike.
Komponenti në rrjedhën e poshtme të kaskadës së transduksionit të sinjalit GPCR përfaqësohet nga proteina G. Janë gjetur rreth 20 proteina të ndryshme G, në radhë të parë duhet përmendur G s dhe C, të cilat përkatësisht stimulojnë dhe frenojnë adenilate ciklazën; Gq, duke aktivizuar fosfolipazën C; G-proteinat e qelizave shqisore: fotoreceptor - G t (transducin), nuhatës - G o nëse dhe shijues - G g.
Proteinat C janë heterotrimerë që përbëhen nga tre lloje nënnjësi: ct, (S dhe y), por në kushte natyrore dy nënnjësitë e fundit funksionojnë si një kompleks i vetëm Ru. Karakteristika më e rëndësishme e proteinave C është prania e a qendra e lidhjes guani në një nën-njësi të tyre - nukleotidet: GDP dhe GTP (Figura 139, 145 nëse GTP është e lidhur me proteinën C, atëherë kjo korrespondon me gjendjen e saj të aktivizuar nëse GDP është e pranishme në qendrën e lidhjes së nukleotideve). atëherë kjo formë i përgjigjet gjendjes joaktive të proteinës (Figura 79).
Ngjarja qendrore në transmetimin e një sinjali nga receptori, i cili ndikohet nga sinjali primar, te proteina G është se receptori i aktivizuar katalizon shkëmbimin e GDP të lidhur me proteinën G për GTP të pranishme në mjedis. Ky shkëmbim GDP/GTP në proteinën G shoqërohet me ndarjen e molekulës trimerike të proteinës G në dy nënnjësi funksionale: nënnjësia α që përmban GTP dhe kompleksin Py (Figurat 139, 145).
Më pas, një nga këto nënnjësi funksionale, e cila varet nga lloji i sistemit të sinjalizimit, ndërvepron me proteinën efektore të përfaqësuar nga një enzimë ose kanal jon. Si pasojë, aktiviteti i tyre katalitik ose përçueshmëria jonike ndryshon në përputhje me rrethanat, gjë që, nga ana tjetër, çon në një ndryshim në përqendrimin citoplazmatik të mesazherit sekondar (ose jonit) dhe, në fund të fundit, fillon një ose një përgjigje qelizore.
Proteinat efektore në sistemet sinjalizuese si GPCR-E G-proteina-E proteina efektore mund të jenë adenilate ciklaza, e cila katalizon sintezën e cAMP nga ATP; fosfolipaza C, e cila hidrolizon fosfatidilinozitolin për të formuar DAT dhe 1P3; fosfodiesteraza, e cila zbërthen cGMP në GMP; disa lloje të kanaleve të kaliumit dhe kalciumit.
Është e rëndësishme që gjatë transmetimit të sinjalit në kaskadën e receptorit të proteinës G-proteinë-efektor, sinjali i jashtëm origjinal mund të përforcohet (përforcohet) shumë herë. Kjo ndodh për faktin se një molekulë e receptorit, ndërsa është në gjendje të aktivizuar (R*), arrin të shndërrojë disa molekula të proteinës G në formën e aktivizuar (G*).
Për shembull, në kaskadën vizuale, rodopsina^C^ecGMP-fosfodiesteraza për secilën molekulë R* mund të prodhojë disa qindra apo edhe mijëra molekula Gt*, që do të thotë se në fazën e parë të kaskadës 7?*-»G* faktori i amplifikimit të sinjalit të jashtëm është 10 2 -10 3 . Megjithëse në fazën tjetër të kaskadës (proteina C*^efektore) secila molekulë G* ndërvepron vetëm me një molekulë të proteinës efektore, sinjali këtu gjithashtu përforcohet, pasi për secilën molekulë të G* dhe, në përputhje me rrethanat, të aktivizuar. proteina efektore, shumë prej tyre shfaqen (ose zhduken) në molekulat e dërguara sekondare të citoplazmës. Kështu, në kaskadën vizuale në fazën e saj të dytë, një molekulë e fosfodiesterazës së aktivizuar cGMP është në gjendje të zbërthejë deri në 3000 molekula cGMP në sekondë, e cila shërben si një lajmëtar dytësor në qelizat fotoreceptore.
Fitimi total i kaskadës është i barabartë me produktin e fitimeve në të gjitha fazat e kaskadës. Koeficienti i amplifikimit të sinjalit kur kalon nëpër kaskadë mund të arrijë vlera shumë të larta: në qelizat vizuale kjo vlerë është e rendit 10 5 -10 6 .
Ndërprerja e stimulit të jashtëm shoqërohet me fikjen e të gjithë komponentëve të sistemit të sinjalizimit. Në nivelin e receptorit, kjo arrihet, së pari, si rezultat i shkëputjes së mesazherit parësor nga kompleksi me GPCR, dhe së dyti, nga fosforilimi i receptorëve nën veprimin e proteinave kinazave speciale dhe lidhja pasuese me një receptor të modifikuar të një specialiteti. proteina (për shembull, P-arrestin).
Proteinat G kanë aktivitet GTPase, domethënë aftësinë për të hidrolizuar GTP të lidhur me to në GDP, gjë që siguron vetëpërjashtimin e tyre, domethënë kalimin G-GTP-e G-GDP. Meqenëse gjendja e aktivizimit të proteinës efektore (on-off) varet drejtpërdrejt nga gjendja e proteinës G, ky kalim nënkupton edhe fikjen e proteinës efektore dhe, rrjedhimisht, ndërprerjen e sintezës (hidrolizës) të proteinës. lajmëtari sekondar ose mbyllja e kanalit jonik.
Dhe së fundi, në mënyrë që të përfundojë kalimi i qelizës në gjendjen e saj origjinale (para stimulit të jashtëm), mekanizma të veçantë rivendosin nivelin fillestar të lajmëtarit sekondar ose kationit në citoplazmën e tij. Për shembull, cAMP, përqendrimi citoplazmatik i të cilit rritet gjatë transmetimit të sinjalit në kaskadën e receptorit P-adrenergjik C5-protein-adenilate ciklazë, hidrolizohet më pas nga cAMP fosfodiesteraza në AMP jo-ciklike (lineare), e cila nuk i posedon vetitë të një të dërguari dytësor.
TRANSMETIMI I SINJALIT TË NDËRTEMBRANËS. Një veti e rëndësishme e membranave është aftësia për të perceptuar dhe transmetuar sinjale nga mjedisi i jashtëm në qelizë. "Njohja" e molekulave sinjalizuese kryhet me ndihmën e proteinave receptore të ndërtuara në membranën qelizore të qelizave të synuara ose të vendosura në qelizë.
Nëse sinjali perceptohet nga receptorët e membranës, atëherë skema e transmetimit të informacionit mund të përfaqësohet si më poshtë:
ndërveprimi i receptorit me një molekulë sinjalizuese (lajmëtari kryesor);
aktivizimi i enzimës së membranës përgjegjëse për formimin e mesazherit të dytë;
formimi i cAMP messenger sekondar, cGMP, IP3, DAT ose Ca 2+;
aktivizimi nga ndërmjetësuesit e proteinave specifike, kryesisht protein-kinazave, të cilat, nga ana tjetër, me anë të enzimave fosforiluese, ndikojnë në aktivitetin e proceseve ndërqelizore.
Ekzistojnë disa mekanizma për transferimin e informacionit transmembranor: duke përdorur sistemin e ciklazës adenilate, sistemin e fosfatit inositol, receptorët katalitikë, receptorët citoplazmikë ose bërthamorë.
Organizimi strukturor dhe funksional i proteinave G
Proteinat G (proteinat që lidhen me GTP) janë ndërmjetës universalë në transmetimin e sinjaleve nga receptorët në enzimat e membranës qelizore që katalizojnë formimin e lajmëtarëve të dytë të sinjalit hormonal. Proteinat G janë oligomere që përbëhen nga nënnjësi α, β dhe γ.
Çdo nën-njësi α në proteinën G ka qendra specifike:
lidhja e GTP ose GDP;
ndërveprimet me receptorin;
lidhje me nënnjësitë βγ;
fosforilimi nga proteina kinaza C;
ndërveprim me enzimën adenilate ciklazë ose fosfolipazë C.
Strukturës së proteinave G i mungojnë domenet α-spiral, që shtrihen në membranë. Proteinat G i përkasin grupit të proteinave "të ankoruara".
Rregullimi i aktivitetit të proteinës G
Ekziston një formë joaktive e proteinës G - kompleksi αβγ-GDP dhe forma e aktivizuar αβγ-GTP. Aktivizimi i proteinës G ndodh pas ndërveprimit me kompleksin aktivizues-receptor, një ndryshim në konformacionin e proteinës G redukton afinitetin e nën-njësisë α për molekulën GDP dhe e rrit atë për GTP. Zëvendësimi i GDP me GTP në zonën aktive të proteinës G prish komplementaritetin midis nën-njësive α-GTP dhe βγ. Receptori i lidhur me molekulën sinjalizuese mund të aktivizojë një numër të madh molekulash të proteinës G, duke siguruar kështu amplifikimin e sinjalit jashtëqelizor në këtë fazë.
A-nënnjësia e proteinës G e aktivizuar (α-GTP) ndërvepron me një proteinë specifike të membranës qelizore dhe modifikon aktivitetin e saj. Proteina të tilla mund të jenë enzimat adenilate ciklazë, fosfolipaza C, cGMP fosfodiesteraza, kanalet Na+, kanalet K+.
Oriz. 5-35. Cikli i funksionimit të proteinës G. R s - receptor; G - hormon; AC - ciklaza adenilate.
Faza tjetër e ciklit të funksionimit të proteinës G është defosforilimi i GTP i lidhur me nën-njësinë α, dhe enzima që katalizon këtë reaksion është vetë nën-njësia α.
Defosforilimi rezulton në formimin e një kompleksi α-GDP, i cili nuk është komplementar me një proteinë specifike të membranës (për shembull, adenilate ciklaza), por ka një afinitet të lartë për protomerët py. Proteina G kthehet në formën e saj joaktive, αβγ-GDP. Me aktivizimin e mëvonshëm të receptorit dhe zëvendësimin e molekulës GDP me GTP, cikli përsëritet përsëri. Kështu, nën-njësitë α të proteinave G lëvizin për të bartur një sinjal stimulues ose frenues nga një receptor që aktivizohet nga një lajmëtar primar (p.sh., një hormon) në një enzimë që katalizon formimin e një lajmëtari dytësor.
Disa forma të protein kinazave mund të fosforilojnë nën-njësitë α të proteinave G. Nën-njësia α e fosforiluar nuk është komplementare me një proteinë specifike të membranës, si adenilate ciklaza ose fosfolipaza C, dhe për këtë arsye nuk mund të marrë pjesë në transduksionin e sinjalit.
Adenilat ciklaza
Enzima adenilate ciklazë, e cila katalizon shndërrimin e ATP në cAMP, është një enzimë kyçe në sistemin e transduksionit të sinjalit të adenilate ciklazës. Enzima i përket grupit të proteinave integrale të membranës qelizore, ajo ka 12 domene transmembranore. Fragmentet jashtëqelizore të adenilate ciklazës janë glikoziluar. Domenet citoplazmike të adenilate ciklazës kanë dy qendra katalitike përgjegjëse për formimin e cAMP, një lajmëtar i dytë i përfshirë në rregullimin e aktivitetit të enzimës së proteinës kinazës A.
Aktiviteti i adenilate ciklazës ndikohet si nga rregullatorët jashtëqelizor ashtu edhe nga brendaqelizor. Rregullatorët jashtëqelizorë (hormonet, eikozanoidet, aminat biogjene) e kryejnë rregullimin nëpërmjet receptorëve të veçantë, të cilët, duke përdorur nën-njësitë α të proteinave G, transmetojnë sinjale për të adenilat ciklazën. Nën-njësia α s (stimuluese), kur ndërvepron me adenilate ciklazën, aktivizon enzimën, nën-njësia α (frenuese) frenon enzimën. Nga 8 izoformat e studiuara të adenilate ciklazës, 4 janë të varura nga Ca2+ (aktivizohen nga Ca2+). Rregullimi i adenilate ciklazës nga kalciumi ndërqelizor i lejon qelizës të integrojë aktivitetet e dy lajmëtarëve kryesorë të dytë cAMP dhe Ca 2+.
Sistemi adenilate ciklaze
Me pjesëmarrjen e sistemit adenilate ciklazë, realizohen efektet e qindra molekulave sinjalizuese të natyrës së ndryshme - hormoneve, neurotransmetuesve, eikozanoideve.
Funksionimi i sistemit të sinjalizimit transmembranor sigurohet nga proteinat: Rs-receptori i molekulës së sinjalit, i cili aktivizon adenilate ciklazën dhe R i-receptorin e molekulës së sinjalit, i cili frenon adenilate ciklazën; Proteinat Gs-stimuluese dhe Gj-adenilate ciklaze; enzimat adenilate ciklaza (AC) dhe proteina kinaza A (PKA).
Sekuenca e ngjarjeve që çojnë në aktivizimin e adenilate ciklazës:
lidhja e një aktivizuesi të sistemit adenilate ciklazë, për shembull, hormoni (G) me receptorin (R s), çon në një ndryshim në konformimin e receptorit dhe një rritje të afinitetit të tij për proteinën G s.
Si rezultat, formohet kompleksi [G][R][O-GDP];
shtimi i [G][R] në G-GDP redukton afinitetin e nën-njësisë α të proteinës G s për GDP dhe rrit afinitetin për GTP. GDP zëvendësohet me GTP;
kjo shkakton shpërbërjen e kompleksit.
Nën-njësia α e ndarë, e lidhur me një molekulë GTP, ka një afinitet për adenilate ciklazën:
[G][R] → [G][R] + α-GTP + βγ;
Ndryshimet konformacionale në kompleksin [α-GTP][AC] stimulojnë një rritje të aktivitetit të GTP-fosfatazës së nën-njësisë α. Ndodh reaksioni i defosforilimit të GTP dhe një nga produktet e reaksionit, fosfati inorganik (Pi), ndahet nga nën-njësia α, dhe kompleksi [α-GDP] mbahet; shkalla e hidrolizës përcakton kohën e transmetimit të sinjalit;
formimi i një molekule GDP në qendrën aktive të nën-njësisë α redukton afinitetin e saj për adenilate ciklazën, por rrit afinitetin e saj për nën-njësitë βγ.
Proteina G s kthehet në formën e saj joaktive;

Sistemi adenilate ciklaze
nëse receptori është i lidhur me një aktivizues, për shembull një hormon, cikli i funksionimit të proteinës G s përsëritet.
Komponenti më i rëndësishëm ndërqelizor i kaskadave sinjalizuese janë proteinat G. Aktualisht, njihen rreth 20 proteina të ndryshme G. Për shembull, Gs dhe Gi stimulojnë dhe frenojnë adenilate ciklazën, përkatësisht; Gq aktivizon fosfolipazën C. Ndër proteinat G të qelizave shqisore mund të veçojmë: fotoreceptor - Gt (transducin), nuhatës - Golf dhe shijues - Gg.
Për sa i përket strukturës së tyre, proteinat G janë heterotrimerë që përbëhen nga tre lloje nënnjësi: a (alfa), b (beta) dhe g (gama), megjithatë, në kushte vendase, nënnjësitë beta dhe gama funksionojnë si një kompleks i vetëm. Një tipar i përbashkët strukturor i proteinave G është prania e shtatë helikave alfa transmembranore. Karakteristika më e rëndësishme e proteinave G është prania në nën-njësinë e tyre α të një qendre lidhëse për nukleotidet guanil: GDP (guanisidin difosfat) dhe GTP (guanisidine trifosfat). Nëse GTP është i lidhur me një proteinë G, atëherë kjo korrespondon me gjendjen e saj të aktivizuar (G-GTP) ose, përndryshe, proteina G është në një pozicion të aktivizuar. Nëse GDP është i pranishëm në vendin e lidhjes së nukleotideve, atëherë kjo formë (G-GDP) korrespondon me gjendjen "off". Pika kyçe në transmetimin e sinjalit nga receptori (i cili u ndikua nga sinjali primar) në proteinën G është katalizimi nga receptori i aktivizuar i shkëmbimit të GDP të lidhur me proteinën G në GTP të pranishme në mjedis (GDP / GTP shkëmbimi në proteinën G). Receptorët transmembranorë sigurojnë funksionet themelore jetësore të qelizës: sinjalizimin, transportin, mbrojtjen. Studimi i mekanizmit të veprimit të komponimeve të ndryshme biologjikisht aktive, duke përfshirë ato antivirale dhe antibakteriale, ka treguar se objektivat më specifikë si për komponimet medicinale ashtu edhe për ato toksike (helmet) janë receptorët qelizor të njerëzve dhe mikroorganizmat patogjenë. Një pjesë e konsiderueshme e receptorëve transmembranorë janë Rreth gjysma e të gjitha barnave të njohura aktualisht veprojnë në mënyrë specifike në GPCR. Nga të gjitha llojet e receptorëve të sipërfaqes qelizore, GPCR-të janë më universalët. Këta receptorë lidhin një gamë të gjerë molekulash, nga neurotransmetuesit e vegjël deri te proteinat e mëdha. GPCR-të përfshihen pothuajse në të gjitha proceset jetësore.
Diversiteti i sinjaleve të transmetuara nga GPCR-të sigurohet nga bashkimi funksional i GPCR-ve të ndryshëm me njëri-tjetrin. Pra, është e qartë se mekanizmi më universal i ndikimit të përbërjeve toksike dhe medicinale në qelizë realizohet nëpërmjet efektit në aparatin receptor të qelizës, duke ndryshuar konformacionin e tyre ose karakteristikat kryesore të lidhjes ligand-receptor, specifikën e tyre dhe kthyeshmëria.

Në Moskë, si në të gjithë Rusinë, jetojnë njerëz të shumë feve. Mes banorëve të kryeqytetit ka edhe protestantë. Ata nuk janë...

Milicia Zemstvo e 1812 Gdhendje nga Ukhtomsky bazuar në pikturën e Luchaninov "Babai bekon djalin e tij për milicinë" Vitet...
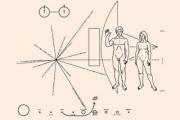
Ideja për të komunikuar për veten me vëllezërit tanë jashtëtokësorë në mendje e vizitoi njerëzimin disa shekuj më parë. gjermane...

Tema: Natyra e atdheut. Qëllimi: të zgjerohen njohuritë e nxënësve për florën dhe faunën e vendlindjes së tyre.

Dashuria... Zë, ndoshta, një nga vendet dominuese në veprën e çdo artisti. Asnjë përjashtim - ...

Dhelpra, pasi nuk e kishte parë kurrë Leon, pasi e takoi atë, mezi mbeti gjallë për shkak të pasioneve të saj, tani, pak më vonë, ajo u përball përsëri me Leon, por ...

Kultura është tërësia e të gjitha llojeve të veprimtarive transformuese të njeriut dhe shoqërisë, si dhe rezultatet...
Lista e shërbimeve të ofruara. Lista e adresave (shtëpive) me të cilat është lidhur kontrata e shërbimit. Kopjet e kontratave në...

Afrika është kontinenti i dytë më i madh në botë. Afrika është e famshme për klimën e saj të nxehtë...

Unë vetëm do të përkthejë saktë!

Që nga viti 1980, ai punoi në shoqatën industriale eksperimentale "Luch", zyra përfaqësuese e së cilës drejtohej ...

Genadi Yuryevich Semigin (lindur më 23 mars) - burrë shteti dhe politik rus, zëvendës...
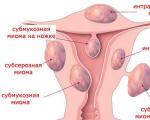
Fibroidet e mitrës, edhe pse një tumor beninj, përbëjnë një rrezik për shëndetin riprodhues...
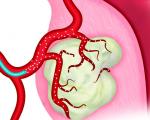
Fibroidet e mitrës janë një neoplazi beninje dhe zakonisht nuk janë kërcënuese për jetën, por kur ato janë të mëdha...

Milicia Zemstvo e vitit 1812 Gdhendje nga Ukhtomsky bazuar në pikturën e Luchaninovit "Babai duke bekuar djalin e tij në...
Ideja për të komunikuar për veten me vëllezërit tanë jashtëtokësorë në mendje e vizitoi njerëzimin disa shekuj më parë.