What is possible and not possible during the Nativity Fast?
In 2018, the Nativity Fast will begin on November 28. During this period, Orthodox believers prepare to celebrate Christmas...
biology Darwin heredity
The modern theory of evolution is based on Darwin's theory, which is why it is called neo-Darwinism. Darwin's main achievement was the establishment of the mechanism of evolution, which consists of natural selection of organisms most adapted to external conditions and the gradual accumulation of acquired characteristics.
The basis of the entire system of modern evolutionary biology is the synthetic theory of evolution, the fundamental provisions of which were laid down by the works of S.S. Chetverikov, R. Fisher, S. Wright, J. Haldane, N.P. Dubinin, Timofeev-Resovsky and others.
The elementary cell of the synthetic theory of evolution is a population - a collection of individuals of the same species that occupies a certain space for a long time and reproduces itself over a large number of generations. The elementary unit of heredity is the gene. Hereditary change of a population in any particular direction is carried out under the influence of such evolutionary factors as the mutation process, population waves, isolation, and natural selection.
Thus, in the synthetic theory of evolution, it is not ontogeny that comes to the fore - the set of transformations occurring in the body from birth to the end of life, i.e. individual development of an organism, but the development of populations.
Structurally, the synthetic theory of evolution consists of theories of micro - and macro - evolution. The theory of microevolution studies irreversible transformations of the tenetic-ecological structure of a population, which can lead to the formation of a new species. The theory of macroevolution studies the origin of supraspecific taxa, the main directions and patterns of development of life on Earth as a whole, including the origin of life and the origin of man. The changes that are studied within the framework of microevolution are accessible to a direct observer, while macroevolution occurs over a long historical period of time and therefore its process can only be reconstructed in hindsight.
The process of evolution was considered slow, mutant genes arose rarely and even more rarely turned out to be more favorable than existing ones. Now many evolutionists believe that in some species evolution occurs according to the “punctuated equilibrium” type, i.e. For a long time, species do not change or the frequencies of different genes remain near a certain equilibrium position determined by general selective factors. Then some sudden change in the environment or a major genetic mutation occurs, changing the gene pool and in a few thousand years a new species with its own genetic balance will appear. Mutations are elementary evolutionary material, but mutations themselves do not ensure evolution, because occur in different directions and can lead to the destruction of what has been acquired. The sources of variability are random genes or chromosomal mutations.
Modern evolutionary theory is often called "synthetic theory of evolution". It includes data from genetics, paleontology, molecular biology, and ecology. N. Green, W. Stause, and D. Taylor define neo-Darwinism as “the theory of organic evolution through natural selection of genetically determined traits.” Biochemists carried out Darwinian evolution at the molecular level. Populations of macromolecules can be forced to develop in the desired direction through cycles of selection, amplification and mutation.
At the core synthetic theory of evolution (STE), or evolutionary synthesis, lies the same Darwinism. However, it is supplemented by information from other biological sciences, primarily genetics, as well as ecology, molecular biology, systematics, etc.
Many features and patterns of the evolutionary process described by Darwin could not be fully explained by him due to the insufficient development of science at that time. This provided the basis for justified criticism of evolutionary theory. For example, Jenkin noticed that any change that arose in one individual would gradually dissolve as a result of crossing and, therefore, could not take hold as a result of the struggle for existence (the so-called “Jenkin's nightmare”). Darwin and his followers could not give counterarguments, since they had little idea of the discrete nature of heredity, although discovered by Mendel in the 60s of the 19th century, but recognized in science only at the beginning of the 20th century.
Discoveries in the field of genetics, molecular biology, the study of populations and the coming to understand how and why a population is the unit of evolution have led to the fact that evolutionary theory has ceased to be something like a hypothesis, but has been explained and largely proven. The mechanisms of evolution were revealed more deeply, the reasons for changes in the gene pool of the population were described, the role of DNA as the material basis of heredity and variability was discovered, and much more.
In the synthetic theory of evolution, the population genetic approach occupies an important place. Population genetics studies how the driving forces of evolution influence changes in allele frequencies in populations, the spatial structure of populations, and explains speciation.
In the 30s of the 20th century, the works of Fisher, Haldane and Dobzhansky showed the relationship between the degree of genetic variability of a population and the rate of its evolution.
In 1942, Huxley proposed the concept of “evolutionary synthesis,” and in 1949 Simpson used the term “synthetic theory of evolution.”
Evolution in STE is considered as a process of gradual change in the gene pool of populations (the frequencies of alleles of different genes change, new alleles and genes appear, others disappear).
According to the synthetic theory of evolution the elementary unit of evolution is the population.
Under elementary evolutionary factors STE and population genetics understand processes that change the set and frequency of alleles. Select a row main evolutionary factors(slightly different in different sources):
Natural selection in the synthetic theory of evolution it is considered as the most important factor in the evolutionary process. As a result of the action of natural selection, the genotypes most adapted to a given environment are reproduced to a greater extent. Natural selection can be directed both against certain alleles and certain genotypes (combinations of alleles and genes).
Struggle for existence. Darwin considered it the main factor of evolution, and natural selection was already a consequence of the struggle for existence.
Mutation process leads to the emergence of new genetic material (new alleles of genes or even new genes). Although mutations occur rarely and are most often harmful, they largely serve as material for the action of natural selection.
Gene flow is a change in allele frequencies in a population as a result of migration of individuals. In addition, gene flow results in the exchange of genes between different populations, which reduces the likelihood of them diverging through the process of speciation.
Genetic drift implies random changes in allele frequencies and is associated with sampling error, which is different from gene flow. Genetic drift in the “founder effect” involves the emergence of a new population from a small number of migrated individuals of another population. These individuals do not carry the entire gene pool of the original population, but only a part of the alleles. In the future, with reproduction and increase in number, the gene pool of the new population will differ from the original one. Another type of genetic drift is the “bottleneck effect,” when the population size sharply decreases as a result of unfavorable conditions.
Insulation- the emergence of barriers between populations that prevent the crossing of individuals and the exchange of genes. As a result, each population can follow its own evolutionary path.
The synthetic theory of evolution shows how heterozygotes (carrying usually harmful recessive mutations) serve as a potential source of the evolutionary process. Recessive alleles are almost never completely eliminated from the population, and when their numbers are small, they remain in heterozygotes.
STE pays great attention to recombination of genetic material. It is often considered as a secondary evolutionary factor, arising on the basis of the above listed primary evolutionary factors that create variability in individual genes, and gene recombination can be considered as a secondary process.
Recombination produces a large variety of genotypes in a population even with a small number of mutations. That is, with an insignificant level of diversity in alleles, a significant level of diversity in genotypes is observed (since the genotype consists of a huge number of genes).
Anyway mutational and recombination variability provide material for natural selection.
As a result of natural selection, adaptations to the environment appear in populations and species, speciation occurs (at the level of microevolution), and the emergence of larger taxa (at the level of macroevolution).
Synthetic theory of evolution. Elementary factors of evolution. Research by S.S. Chetverikova. The role of evolutionary theory in the formation of the modern natural science picture of the world.
When we speak from, we always associate this concept with the name. A scientist lived in the 19th century, and debates about evolution (in particular) continue to this day.
In addition, genetics, which also originated around the 19th century, is now moving forward by leaps and bounds. Therefore, it would be logical to combine existing knowledge and accumulated factual material into a new theory of evolution - a more modern one.
Synthetic theory of evolution- modern Darwinism - arose in the early 40s of the XX century. It is a doctrine of the evolution of the organic world, developed on the basis of modern and classical data. The term " synthetic" comes from the title of the book by the famous English evolutionist J. Huxley " Evolution: a modern synthesis"(1942). Many scientists contributed to the development of the synthetic theory of evolution.
As in the classical The main provisions of the synthetic theory of evolution were put forward:
The man who first combined the knowledge of genetics and Darwin's evolutionary theory is S.S. Chetverikov.
Like Darwin, Chetverikov understood the importance of factual material - he experimentally studied the inheritance of properties in animal populations under natural conditions.
For 9 years he worked as the scientific director of the genetics department of the Institute of Experimental Biology, one of the largest institutes of that time.
In his works, he showed that there is no contradiction between genetic data and evolutionary theory. On the contrary, genetic data should form the basis of the doctrine of variability and become the key to understanding the process of evolution. Chetverikov proved that mutations in natural animal populations do not disappear, but can accumulate in a latent (heterozygous) state and provide material for variability and natural selection.
But, as is known from, the life of our scientists was not easy. At the end of the 30s he was convicted and exiled for 6 years. After exile, he continued his work, but not in the capital, but in the city of Gorky... but also as the head of the department of genetics at Gorky University.
It is he who is considered the founder of the synthetic theory of evolution; it is also sometimes called “neo-Darwinism”, paying tribute to the man who was the first to build the available facts into a clear, logically based chain of theory.
From Igor Garin's book "Darwin". Notes and citations appear in the text of the book.
We cannot categorize traits into two separate boxes, one labeled "hereditary" and the other labeled "environmental." Everything in the body is the result of the interaction of both.
M. Bates
The first attempt to combine Mendelism and Darwinism was made, as already noted, by Thomas Hunt Morgan: according to his point of view, mutations play a leading role in the origin of species, while selection acts like a sieve, preserving rare beneficial mutations and eliminating all others. Morgan's conclusions received experimental confirmation in an experimental study of microevolutionary processes: selection turned out to be ineffective within a genotypically homogeneous group (“pure line”) and when acting on fluctuating variability, and was effective in a genetically heterogeneous population before pure lines were isolated from it. Later, however, it became clear that selection can also be effective in the Darwinian sense if spontaneous hereditary changes occur within genetically homogeneous material. In the 20s, genetic experiments, conceived to refute Darwinism, increasingly led to the resolution of contradictions in evolutionary theory and to the combination of De Vries mutations and Darwinian variability in the theory of microevolution.
After the rediscovery of Gregor Mendel's laws by H. de Vries, K. Correns and E. Cermak, the most important discovery in genetics was the mathematically proven constancy of the frequency of occurrence of different alleles in a population over a number of generations (Hardy-Weinberg rule). From this rule it followed that evolution is the process of changing the gene pool of populations, depending mainly on the process of mutation and natural selection. If at the beginning of the century genetic mutation was opposed to selectogenesis, then in the 20s, thanks to research in population genetics and the use of mathematical methods of analysis in the theory of evolution (the work of John Haldane, Ronald Fisher and Sewall Wright), a new concept of evolution began to emerge, synthesizing the latest discoveries of population genetics with Darwinism.
The greatest merit of Haldane, Fisher and Wright, whose names are usually mentioned together as the creators of the theoretical basis of population genetics, * was that, having substantiated the “discrete” nature of inheritance, they developed probabilistic and statistical methods for describing the inheritance of traits and selection. In one of his first articles, John Haldane wrote:
A satisfactory theory of natural selection must be quantitative. In order to establish that natural selection is capable of explaining the known facts of evolution, we must show that it not only causes changes in a species, but that it can cause changes in that species at a rate which will account for present and past transmutations.
If Darwin thought in words, then his followers today must think in numbers.
John Haldane proceeded from the fact that natural selection is the selection of large or minor mutations, the nature of which was not yet understood, but the fact of whose existence was already firmly established:
...The fact that they [mutations] do occur is sufficient to make evolution by natural selection possible and intelligible.
According to D. Haldane, selection can act only when mutations supply it with material. Neither mutations themselves, nor selection itself separately can serve as the basis for long-term evolution:
Quantitative analyzes show that natural selection is a reality and that among other objects it selects for Mendelian genes, which are known to be randomly distributed throughout wild populations and follow the laws of chance in their distribution in descendants. In other words, they are the means of producing variation of the kind postulated by Darwin, forming the raw material upon which selection acts.
D. Haldane was most concerned with the problems of the pace of the evolutionary process: the intensity of extinction, the growth of populations, the rate of selection, an increase or decrease in the concentration of individual genes in a population, etc. One of the main results of his calculations is the “incredibly low rate of evolution”: “.. "In most species no noticeable changes can be expected within 10,000 years." Although John Haldane showed that the slow rate of mutation characteristic of most genes is not a limiting factor in evolution, in his recent work he drew attention to the fact that for mutations to spread, the rate of their occurrence must be much higher than usually observed in nature.
For the revival of the theory of natural selection, Ronald Fisher's book, The Genetic Theory of Natural Selection, was of great importance. which shows that genetic variation depends on population size. Although mutations are often less adaptable and usually recessive, they constantly arise again and again, albeit with low frequency (this is due to the slowness of the evolutionary process). However, in large enough populations the prospect of adaptive change and the survival of mutants increases dramatically, and numerous species have a marked potential for variation sufficient to allow evolution. According to R. Fisher, the probability of the emergence of a new species through a single macro-leap is negligible: for a complex organism, any sudden change will be unfavorable. The main mechanism of evolution is therefore small, but proceeding at a certain rate of saltation.
P. Mahalanobis:
Like Clarke Maxwell, who translated Faraday's concepts into mathematical language and developed the electromagnetic theory of light, Fisher quantified Darwin's views and constructed a statistical theory of evolution.
Unlike Fisher, who believed that natural selection operates most effectively in large populations, where a large variety in the combination of genes is possible, Wright, on the contrary, concluded that natural selection is most active in small populations, where the “genetic drift” he discovered is most likely. “Genetic drift” (N.P. Dubinin and D.D. Romashov called it a “genetic-automatic process”) is a natural variation in the gene pool of a population without the participation of natural selection. In small and especially isolated populations, some genes can be completely lost, while others can be acquired without any selective significance of the corresponding traits. According to Wright, “genetic drift” can lead to the formation of new forms without selection and even in spite of it.
An important difference between Wright’s ideas and his predecessors was the connection of evolutionary transformations not with the interaction of individual genes, but with their systems. This idea subsequently prevailed.
Proposed in 1908–1909 by the English mathematician G. G. Hardy and the German physician V. R. Weinberg, the Hardy-Weinberg rule, which explains the arbitrarily long preservation of two alleles of one gene in a population - dominant A and recessive a, is valid only if the following restrictions are met: 1) an infinitely large population ; 2) unlimited crossing options - absolute panmixia; 3) absence of mutation process; 4) no differences in the selective value of different alleles; 5) the absence of overlapping generations in time (which is characteristic, for example, of many salmon fish, cicadas, May beetles, but is unusual for the vast majority of other groups).
Failure to meet these ideal conditions leads, even in the case of equal selective value of the A and a alleles, to the random displacement of one allele by another, which ultimately leads to the fact that some populations may eventually turn out to be monomorphic for the A allele, and others for the a allele. This random displacement of one selection-neutral (i.e., with equal probability of survival under the influence of similar pressure of natural selection) allele by another was called “genetic drift.”
Almost simultaneously and independently of Fischer and Wright in the USSR in 1931–1932. Dubinin and Romashov, who proposed the term “genetic-automatic processes” for this phenomenon, came to the same conclusions. It should be noted that the mathematical model of random or stochastic processes leading to the divergence of populations due to genetic drift (genetic-automatic processes) follows from the so-called “Markov chains” or “Markov processes”, the theory of which was developed in abstract form in 1907 Russian mathematician A. A. Markov Sr.
An important contribution to the development of evolutionary theory was the work of O. S. Chetverikov, confirming the genetic heterogeneity of the natural population, which serves as the genetic basis of the evolutionary process. Chetverikov established that the mutation process occurs in natural populations, and mutations that increase viability are rarely encountered. As a species ages, an increasing number of mutations accumulate in it, expanding the characteristics of the species. Genotypic variation increases when a large species breaks up into a number of small, isolated colonies.
Chetverikov’s research confirmed that natural populations, “like sponges,” are saturated with various mutations, that is, they represent a very heterogeneous mixture of different genotypes. This change in the genotypic composition of the population is the basis of its evolution. Adaptive evolution without isolation can lead to complete transformation of a species, but cannot lead to divergence. Intraspecific differentiation requires isolation. Selection selects not only a single gene that controls a single trait, but, due to the multiplicity of gene action, influences the genotypic environment.
Chetverikov also belongs to the fundamental idea of non-adaptive evolution, according to which the appearance of new characters may precede their use in the subsequent development of the organism:
We have no reason to deny the possibility of non-adaptive evolution. In many cases, it can be assumed that the existing adaptive differences between the basic forms were not the reason for the divergence of the latter, but, on the contrary, the specific nature of these adaptive reasons is a consequence of the earlier separation of forms.
Based on the Hardy-Weinberg rule, Chetverikov formulated the idea that the mechanism of free crossing ensures the balance of a given community, stabilizing the number of its components:
Its direct antagonist in this regard is natural selection (and selection in general in any of its forms). If free crossing stabilizes the community, then selection, on the contrary, constantly takes it out of equilibrium, and if in this sense we can call free crossing a conservative principle, then selection is undoubtedly an evolutionary principle, continuously leading to a change in the species.
In other words, there are two differently directed forces: stabilization of the community by crossing and disruption of equilibrium by the action of selection.
From the point of view of the development of the theory of natural selection, the most important thing was that Chetverikov considered natural selection as the cause of adaptive evolution, that is, a factor that is responsible for increasing the fitness of species. Chetverikov's following conclusions were of great importance. Firstly, he believed, as a result of free crossing and selection, under the conditions of Mendelian heredity, even small “improvements” in individuals have a chance of spreading among many individuals of the species. Thus, Mendelism, as Chetverikov emphasized, becomes a true and extremely important and unexpected ally of Darwin's theory. The second conclusion was that the selection process itself cannot lead to the formation of two species, that is, the splitting of a species. The latter becomes possible only in cases where some type of insulation appears. Chetverikov substantiated all these conclusions with specific, mainly entomological, observations.
Chetverikov has another idea - “waves of life,” a periodic increase or decrease in population numbers. As it turned out later, these “ebbs and flows” associated with imbalances are not an accident and play a certain role in evolution.
The evolutionary significance of “waves of life” and fluctuations of territories occupied by populations is that a process that is random in nature influences the direction and intensity of selection pressure, changing the concentration of different genotypes and mutations in natural populations and being the basis for “testing” new genotypes.
Chetverikov considered population waves and isolation to be the most important driving forces of evolution, in particular, isolation causes and consolidates group differences due to the disruption of panmixia leading to leveling *. Although the presence of “life waves” is currently not questioned by anyone, neither their nature nor their role in evolution has been clearly established. In particular, the question of whether population waves are a self-sufficient factor of evolution or a consequence of known ones continues to be discussed.
In Genetics and the Origin of Species, F. G. Dobzhansky, following Chetverikov, introduced the half-forgotten Hardy-Weinberg rule into wide scientific circulation and developed an important population genetics, the concept of “isolating mechanisms of evolution” - barriers separating the gene pools of species.
The foundations of population genetics, laid by S. S. Chetverikov at the Institute of Experimental Biology, were further developed in the works of students and followers, who primarily include I. V. and E. A. Timofeev-Resovskikh, B. L. Astaurov, N. P. Dubinin, S. M. Gershenzon, F. G. Dobzhansky and others.
The synthesis of genetics and selectogenesis was also facilitated by the “law of homological series” formulated by N. I. Vavilov, according to which similar hereditary changes occur in species of close origin. Vavilov’s work made it possible for the first time to predict mutations fixed by natural selection. Vavilov also belongs to the concept of a wide polytypic species in botany: like Ya. Lotsi**, Vavilov divided Linneon into a system of Jordanons subordinate to it. Thus, the idea of polytypic species was formulated, which later became widespread, with its characteristic transition to a population way of thinking.
In the mid-20s, direct experiments by G. A. Nadson, G. S. Filippov and G. Meller discovered inherited mutations caused by radiation. Thus, the existence of hereditary variability was proven experimentally. The first experiments on natural selection, carried out by A. Chesnola on mantises *, and then continued in the works of M. M. Belyaev, M. Geller, K. Swinnerton and others, revealed that the most important factor of selection is protective coloration: insects were eaten by birds much more faster if placed on unsuitable backgrounds for their coloring. Insectivorous birds represent a fast-acting factor of natural selection, promoting the development of protective (or warning) coloration in insects. The darkening of color in many species of butterflies living in industrial zones as a result of soot deposits on tree bark (so-called industrial melanism) is also considered to be evidence of the effectiveness of natural selection. In this case, selection is also associated with the destruction of light-colored butterflies by birds and a clear advantage of melanistic forms in industrial areas. In these cases, we are dealing with pure Darwinian selection, not associated with mutations.
In the forties, the development of the synthetic theory of evolution (STE) was associated with the works of J. G. Simpson, F. Dobzhansky, J. Huxley, E. Mayr, B. Rensch. Simpson owns the term “quantum evolution,” which he proposed to explain “evolutionary explosions.” The book Quantum Evolution substantiates the role of saltation as the dominant and most significant process in the formation of large taxa - families, orders and classes, the emergence of which is associated with the emergence of completely new adaptive types, then there are shifts from one adaptive zone to another.
J. G. Simpson:
The entire [geological] record indicates that the emergence of a new adaptive type normally occurs much faster than further processes of progressive adaptation and diversification within this type.
According to Simpson, “the most radical types of quantum evolution probably begin by the random fixation of non-adaptive mutations in very small, isolated populations.”
Based on the idea of the unity of the mechanisms of micro- and macroevolution, Simpson explained the incompleteness of the geological record by the small number of populations of transitional forms and the high intensity of natural selection under conditions of sudden changes in the environment, which sharply reduced the likelihood of their detection in a fossil state.
According to Simpson, there are three main forms of evolution: fissionable, phyletic and quantum*. Splitting is determined by the differentiation of a population with a large range into local adaptive groups, which are gradually transformed into new subspecies. Split evolution is characterized by its reversibility, at least in the initial stages, as well as lack of direction and low speed of transformations.
Phyletic evolution is directional because it is caused by a change in the adaptive zone and gives rise to a new form of the same taxometric rank, usually leading to the emergence of new genera and families.
Unlike the two indicated forms, quantum evolution is associated with the transition to a new adaptive zone. It occurs at a rapid pace, but spreads to a small part of the population. The incompleteness of the geological record is associated precisely with quantum evolution, in which small and migratory populations, consisting of individuals with rapid succession of generations and living in unstable environmental conditions, have almost no chance of preserving their trace in the fossil state. In other words, “quantum speciation” (Simpson) or “genetic revolutions” (Mayr) are not recorded in the geological record due to the rapidity of change and the small number of transitional forms.
We see that Simpson, without abandoning the idea of selection, abandons the idea of gradualism:
An impartial consideration of many of the phenomena of so-called orthogenesis clearly shows that much of the straightforwardness of evolution arises rather from the tendency towards straightforwardness of the minds of scientists than from a similar tendency in nature.
The latest research has discovered strong intraspecific differentiation and a complex hierarchy of intraspecific units. Along with variation within a population, there is interpopulation variation: each population is genetically different from other populations, and the degree of difference between them varies from negligible to almost interspecific.
E. Mayr:
Geographic variation is a population phenomenon that contributes significantly to understanding the nature of species. It shows the flaw of the typological concept of a species and allows us to conclude that a significant part, if not all of the variability, is adaptive and arises in response to a variety of environmental requirements.
According to population genetics, the main component of speciation is changes in the genetic structure of populations. It is thanks to polytypicity and genetic diversity that geographic speciation under conditions of isolation, predicted by M. Wagner in 1868, is possible. Although geographic isolation, which is an important prerequisite for the formation of animal species, is eroded by sexual intercourse and does not in itself lead to the formation of species, it makes possible the unhindered genetic restructuring of populations, which, in turn, is a prerequisite for the creation of isolating mechanisms.
A more detailed study of mutations made it possible to establish that, being discrete changes in the codes of hereditary information, they themselves are very different in nature and occur at different levels: genes, chromosomes, genomes and extranuclear DNA-containing structures of the cytoplasm. Spontaneous mutations occur with a certain frequency in all organisms and are the main “supplier” of new elementary material in the population. They support a high degree of heterogeneity in natural populations.
It is generally accepted that there are different genetic systems, some of which promote specialization, others - broad general adaptation, and others - the role of pioneers in evolution.
E. Mayr:
In order to achieve maximum evolutionary success, a phyletic lineage must have the ability to switch from one type of development to another. The pioneer of evolution, having made a breakthrough, must develop widely adapted species to ensure his future.
As a result of the geometric progression of reproduction and hereditary changes that determine the inexhaustible genetic diversity of the population, “life pressure” arises, leading to natural selection and subsequent reproduction of only a small number of individuals born. According to Timofeev-Resovsky and co-workers:
An elementary evolutionary phenomenon - a sustainable change in the genotypic composition of a population - arises as a result of the pressure of the mutation process, population waves, isolation and always occurs under the guiding action of natural selection.
In the process of evolution, individual individuals can acquire properties that make effective crossing with other individuals impossible; the emergence of reproductive isolation means the cessation of intraspecific leveling through crossing and ultimately leads to the emergence of a new species as a qualitative stage in the evolutionary process. With the emergence of a new species, interspecific competition arises and interspecific selection begins to operate. All this leads to the amazing diversity and richness of the organic world.
According to STE, not a single gene, taken separately, has a constant selective value: it can either significantly increase fitness or sharply reduce it, depending on the general genetic background (genetic theory of relativity). Selection works only against the background of the harmonious interaction of all genes, trying to bring together those genes that make up a balanced system.
E. Mayr:
The process of incorporating harmoniously cooperating genes into the gene pool is called “integration” or “coadaptation.” The result of such selection can be called epistatic equilibrium. Each gene will be favored by selection for the genetic background in which it is most likely to contribute to increased fitness. Thus, the fitness of a particular gene depends on the entire genetic background and is controlled by this genetic background.
The result of coadaptive selection is a harmoniously integrated complex of genes. The combined effect of these genes can manifest itself at many levels - at the level of the chromosome, nucleus, cell, tissue, organ and the whole organism. For the evolutionist, the nature of the functional mechanisms of physiological interaction is of only secondary interest; he is mainly interested in the viability of the final product - the phenotype.
Phenotype is a by-product of a long history of selection and is therefore highly adapted. The influence of selection is usually expressed in the stabilization or normalization of the phenotype. Because this well-integrated phenotype is adapted to maximize its contribution to fitness, it resists change (inertia, genetic homeostasis) when new selection pressures arise.
The uniqueness of each chromosome, each individual (in dioecious species) and each population determines a huge and unpredictable variety of reactions to the action of selection.
E. Mayr divided all theories of evolution into monistic or single-factor theories, which explain evolutionary changes by the action of one factor - the external environment, internal forces, random events, natural selection, and synthetic ones, which explain evolutionary changes by the action of many factors. Being a supporter of the synthetic theory of evolution, he preferred the “modern synthesis”, which combined spontaneous mutations and mutational restrictions with natural selection and undirected response to environmental changes.
Attempts to explain evolution on the basis of any one factor proved fatal to pre-Darwinian evolutionary theories and most evolutionary theories of the 19th century. Lamarckism with its internal principle of self-improvement, Geoffreyism, which postulates the induction of genetic changes by the external environment, Cuvier's catastrophism, Wagnerian evolution by isolation, De Vries mutationism - all these were attempts to explain evolution, relying on one principle and excluding all others. Even Darwin sometimes made this mistake, but it was he who first made a serious attempt to represent evolutionary events as the consequence of a balance of opposing forces. The modern theory of evolution - the "modern synthesis", as Huxley called it - owes more to Darwin than to any other evolutionist, and is built on the basis of basic Darwinian concepts. However, it included many post-Darwinian provisions. Concepts such as mutation, variation, population, heredity, isolation and species, still very vague in Darwin's day, are now more clearly understood and much more strictly defined.
According to E. Mayr, the development of STE was greatly facilitated by the overcoming of two ideological misconceptions - preformationism and typologism, and it was the rejection of incorrect philosophical premises, and not particular theories of evolution, that had a decisive impact on the development of modern ideas.
Preformationism is a theory of development according to which a transformed individual in miniature is “enclosed” in an egg or sperm and only needs to “unfold” during development. The term "evolution" was borrowed from this concept of unfolding, and this misinterpretation persisted long into the post-Darwinian period. This is probably why Darwin did not use the term “evolution” in his book On the Origin of Species. Transferred from ontogeny to phylogeny *, the term “evolution” denoted the implementation of a certain initially laid down plan. Evolution, according to this point of view, does not cause genuine changes, but consists only in the realization of initially inherent potentials. This was, for example, Agassiz's theory. Some of the orthogenetic and finalist theories of evolution represent the last vestiges of this type of thinking. Mutationism was an extreme form of reaction to these orthogenetic concepts. Modern theory makes a compromise by recognizing that the genotype and phenotype of a given evolutionary line determine the strict limits of its evolutionary potential, without, however, predetermining the path of further evolutionary changes.
The typological concept is another major flawed concept that had to be completely refuted before a coherent theory of evolution could be proposed. The philosophical basis of this form of thinking is Plato's concept of idea (eidos). According to this concept, the observed diversity of the world is no more real than the shadows of some objects on the wall of a cave, as Plato allegorically expressed it. It is the permanent and unchanging “ideas” that underlie the observed diversity that are the only permanent and real ones. Since adherents of this philosophy believe in essences, the philosophy itself is called essentialism, and its adherents are called essentialists (typologists). Most of the great philosophers of the 17th, 18th and 19th centuries. were influenced by this idealistic philosophy of Plato and its modification proposed by Aristotle. The way of thinking inherent in these schools dominated the natural sciences throughout much of the 19th century. The concepts of unchanging entities and the complete isolation of each idea (type) from all others made truly evolutionary thinking almost impossible.
E. Mayr:
Potentially, speciation represents a process of evolutionary rejuvenation, a way to avoid a system of genetic homeostasis that is too rigid. Speciation breaks the integrity of the gene pool, temporarily impoverishing its gene content and inevitably forcing the population to move to slightly different or dramatically different environmental conditions. If this genetic "shake-up" is strong enough, it can start a chain reaction - a genetic revolution. The stronger the genetic changes, the greater the likelihood that the daughter species will be able to penetrate a new ecological niche and exist successfully in it. Thus, a genetic chain reaction can start an evolutionary chain reaction. This process, in its purest form, is most likely to occur in peripheral isolated populations.
Speciation is a risky process. The impoverishment of the gene pool and genetic instability that accompany speciation are much more likely to lead to disaster than to success. However, although most nascent species die, one will not only complete this process, but will also successfully penetrate a new niche or adaptive zone.
The significance of speciation is that it promotes evolutionary experimentation. It creates new units of evolution, particularly those that are important for potential macroevolution. Speciation is a progressive process, not a regressive one.
Mayr considered the worldviews of a typologist and a populationist to be mutually exclusive: for a typologist, eidos and ideas are real, and variability is illusory; for a populationist, a type is an abstraction of averaging and only variability is real. “The replacement of typological thinking by thinking in population terms,” writes E. Mayr, “was probably the greatest conceptual revolution that has ever occurred in biology.”
According to E. Mayr, the leading factor in evolution is selection, not genetic rearrangement:
1. Evolution cannot be considered a primarily genetic phenomenon. Mutations only provide the gene pool with genetic variability; the factor causing evolutionary change is selection.
2. Each trait is usually not the result of the action of a single gene; therefore, a change in a trait does not indicate a mutation of any one gene. In fact, all traits are highly polygenic, and since most genes are pleiotropic*, a change in one trait indicates a greater or lesser restructuring of the genotype.
3. The assumption that genes can be divided into beneficial ones, which are automatically included in the gene pool, and disadvantageous ones, which will be ruthlessly eliminated, is only misleading. The selective value of a given gene is not absolute: in each individual case it is largely determined by external conditions and the epigenetic system in which this gene operates. Consequently, natural selection cannot be represented as a simple arithmetic problem. In addition, it must be assumed that the genes of a certain gene pool, being selected for their coadaptive ability, on average increase fitness more than randomly added genes.
If Darwin had to build the theory of natural selection mainly on the logical consequences of the factors observed in nature in the adaptation of organisms to the conditions of existence, now different forms of selection have been studied in detail, experimentally confirmed and classified - leading and stabilizing (according to Schmalhausen), driving, stabilizing, destabilizing and crushing (according to Simpson and Mather).
When analyzing natural selection, Schmalhausen proceeded not from Darwin's concept of selection of the fittest, but from Morgan's idea of eliminating the least fit. In this case, the forms of natural selection are determined by the forms of elimination.
Thus, the question of the selection role of minor favorable changes completely disappears. We can only talk about the eliminating significance of unfavorable changes. In the process of evolution, there is a free accumulation of not only minor positive, but also indifferent and sometimes even partially harmful mutations. Therefore, the constant indications of anti-Darwinists that species differences are in many cases of an indifferent nature do not in any way contradict Darwin’s understanding of speciation.
Leading (driving) selection is necessary when the conditions of existence change, when it is not the “norm” that gains advantage, but the “deviation.” It leads to a change in species - the emergence of a new norm of reaction and the elimination of the old, less adapted one. Stabilizing selection, on the contrary, manifests itself when conditions are favorable for the “norm” - the development of the most stable and viable phenotype in given environmental conditions, when the elimination of random deviations is accompanied by an increase in the stability of the new “norm”.
An important hidden subtext of the theory of evolution is what is called the principle of convariant reduplication - the desire for self-preservation, which can only be achieved through constant self-renewal. The preservation and evolution of life are possible only with a constant change in its content. In other words, without development, life is doomed to degradation (an example of which at the social stage of its development is its communization).
To summarize, we can say that within the framework of the synthetic theory of evolution, which denies vitalist concepts, we are dealing with a synthesis of genetic concepts and the theory of natural selection. The STE formula can be represented as: “mutation - gene recombination - selection”. The main postulates of STE can be formulated as follows:
- Darwin's small but discrete changes in heredity - mutations - serve as the material for evolution. Mutational variability is random and is not caused by the body's needs or environmental changes.
- The driving factor of evolution is natural selection, which, however, is not the only one. Isolation or genetic drift may also play a significant role. At the same time, genetic drift should not be opposed to the action of selection.
Thus, we can talk about two stages of evolutionary change: genetic variability (through mutations, recombinations and random events), and variability subjected to ordering through natural selection.
The smallest evolving unit of evolution is not an individual, but a population, and the evolution of a population is predominantly divergent in nature, that is, species are formed due to the divergence of characters, so that one taxon can become the ancestor of several daughter ones.
- Although in general evolution is gradual and long-term, transformations through “explosions” and macromutations are allowed, but in any case the mechanisms of micro- and macroevolution coincide.
- Species are polytypic in nature, but at the same time the species itself is a genetically integral and closed system, protected by a complex system of barriers and isolating mechanisms of evolution that prevent the exchange of genes between gene pools of different species.
- The random nature of mutational variability is compatible with its specific direction, determined by the previous course of evolution.
Despite the large number of adherents of STE and the deep scientific study of its individual positions, our assertion that after the creation of STE the crisis of Darwinism was overcome is completely untrue. Both in the 30s (G. Wolff, M. Thoma) and in the 50s (O. Kuhn, J. Klotz, D. Devor, N. Heribert-Nilsson, L. Bunur, A. Rouvier, J. Corle ), and in the 60s (R. Zdanskaya, A. Pirovano, A. Meyer-Abikh) and in subsequent years, fundamental monographs appeared that questioned the compatibility of Mendelism and Darwinism. In particular, N. Heribert-Nilsson wrote:
Mendel's proof of the fundamental constancy of genes makes bridging between species a much more difficult task than Darwin thought.
F. Dobzhansky:
It would be wrong to say that the biological theory of evolution has gained universal acceptance among biologists or even geneticists.
E. Mayr:
...a deeper understanding of the nature of the gene and the mutation process has added almost nothing to the understanding of evolutionary phenomena.
Considering STE as “ultra-Darwinism,” A. A. Lyubishchev considered the claims of the followers of Haldane, Fisher and Wright to create a general theory of evolution untenable due to the construction of the theory as universal and irrefutable. He recognized the role of STE in microevolution, but considered it unacceptable for explaining macroevolution, where its extrapolations are not sufficiently justified. The nature and mechanism of evolutionary processes at different levels of organization should be different. Lyubishchev also considered the disadvantages of STE to be the tendency to explain all evolutionary phenomena by a “leading” factor; denial of psychoid factors; abuse of evidence by contradiction and the law of exclusion of third. Using the terminology of genetics, Lyubishchev believed that STE hardly attracts the results of the latest genetic research, remaining within the conceptual framework of genetics of the 30s. As a supporter of Berg's nomogenesis, Lyubishchev believed that hereditary variability is not chaotic, but directed, and in the course of evolution a limited number of regular discrete possibilities are realized. Goal setting is the inner essence of life. The appearance of similar forms and organs on different branches of the phylogenetic tree is the result of nomogenesis.
Like Darwinism, STE does not explain in any way the fairly common phenomenon of pre-adaptation - the emergence of traits, the need for which arises only in the course of further evolution. No successive mutations or selection can produce adaptations that, at the time of their occurrence, are meaningless and exceed the biologically necessary level. We are dealing with a certain reserve of adaptability, acquired not due to biological necessity, but as if by the presence of a distant goal.
A. B. Georgievsky:
At present, of course, it is quite difficult to explain the amazing mystery of living nature - the development of such states of adaptation that are not required in real life conditions.
Like Darwinism, STE is powerless to explain progressive evolution, ascending to higher forms, including the emergence of the mind and soul. Within the framework of STE, the incentives for increasing complexity are not clear.
STE does not adequately explain either stagnant or explosive evolution. The fundamental question of the interactions of genotypes with the external environment, as a result of which a phenotype arises - the real object of natural selection - still remains controversial. E. Mayr lists a number of issues on which STE representatives have not yet come to a common opinion:
How important are random events in evolution?
How big is the role of hybridization in evolution?
What are the consequences of gene exchange between populations?
What proportion of new mutations are beneficial?
What proportion of genetic variation is due to balanced polymorphism?
How important is phenotypic plasticity?
It should be noted that even among STE adherents, not everyone is satisfied with the state of the theory. J. G. Simpson believed, for example, that STE does not explain a number of significant phenomena in the development of life, in particular, the direction of evolution:
We may really know nothing about the causes of directionality, or these causes may actually be transcendental, but giving names to what we do not know does not alleviate our ignorance, and postulating transcendence always nullifies the inquiry.
Criticizing, in turn, the views of Simpson himself, J. Brow in his work Time and Evolution comes to the conclusion about the existence in the past of “great waves of evolution” that led to the rapid and sudden genetically determined emergence large taxa: phyla, classes, orders and, perhaps, families. The selection factor, if any, was involved only in the elimination of bad genetic material, and not in morphogenesis. Mutation rates in the past could be very high, and the mutations themselves could be directed. Subsequently, as the forces of evolution were exhausted, the speed of evolutionary processes began to decrease, and the entire biosphere began to age:
The variability of organic forms has nothing to do with environmental factors: it depends on spontaneous mutations.
The field of evolution is constantly shrinking, and in the future it will completely stop.
There is no doubt that human activity is a powerful factor in inhibiting evolution and the physical destruction of the diversity of life as such. By the way, according to M. Bates, evolution is a close interweaving of genetics and ecology, and selection is a purely ecological cleansing process.
Currently, there is a fairly widespread opinion that STE is nothing more than “theoretical euphoria” (J.-P. Leman), revealing a “fearful tendency to turn into an illiberal orthodoxy that does not tolerate dissent” (M. Raduik). The resuscitation of Darwinism in STE does not correspond to its real contribution to understanding the development of the biosphere.
F. R. Shram:
Ehrlich and Raven questioned the genetic basis of [STE], namely the recognition of the essential role of genetic variation and genetic drift in explaining the origin of species. They showed that small-area natural populations can retain strong heterogeneity even when gene flow is greatly limited, and this limitation can be considered a common property of wild populations. Although under normal conditions the choice of mating partner for an animal or plant individual is extremely limited, these highly localized populations retain a high level of genetic diversity. Individuals within an animal or plant species typically have little or no freedom to choose a mating partner, yet such local populations retain high genetic diversity. Taking into account these limitations of crossing in natural conditions reveals the incorrectness of the definition of a species based on the concept of a “common gene pool”, based on the idea that in a geographically compact species all the genes of crossing individuals are at the disposal of the species as a whole. Ehrlich and Raven propose to discard these two cornerstones of the synthetic theory: the common gene pool and the genetic mixing that leads to diversity.
Because in STE selection acts randomly on random genetic variations, in synthetic theory evolutionary changes are not defined or directed. In other words, we can say absolutely nothing about where the phylogenetic line and evolution in general will “lead”. Evolution as a process turns out to be only a sliver in the flow of external conditions, which can carry it anywhere *.
There is also a more fundamental objection to STE as a paradigm built on a classical, that is, Newtonian basis, which does not take into account the theory of relativity, quantum mechanics, the uncertainty principle and causality in its modern understanding.
Thus, as F. R. Schram said, modern biology faces an alternative: adapt to the imperfections of the synthetic theory of evolution or look for another basis for creating a paradigm.
The unspoken assumption underlying all theories of evolution is that its laws are eternal and unchanging: three billion years ago, processes on Earth were approximately the same as they are now. But where is the proof that the mechanisms of evolution do not themselves evolve? What is the basis for the assumption that the interaction of evolutionary factors in completely different conditions of the Earth and in unimaginably distant times has remained unchanged?
K. M. Zavadsky:
The uniqueness of the organization determines the uniqueness of development. After all, if, say, the hereditary substance of an organism is not yet even organized into chromosomes, then the factors of evolution of such an organism will manifest themselves differently than in higher animals... On this basis, the point of view has been expressed more than once that Darwinism is correct only for modernity, but completely inapplicable for distant eras.
Nowadays, even such a fundamental concept as the “ladder of life”, or the uniqueness of the hierarchy of living things, is being questioned, and a new paradigm is being developed for the original existence of many “primary” forms that gave rise to different trunks of the phylogenetic tree. According to V.I. Vernadsky, life is initially a complex of complex forms: it immediately arose as a complex system. The problem is not even the direction of evolution, the improvement and complication of life, the problem is the systematic nature of the initial life, which during its natural occurrence contained something more than aperiodic crystals, because aperiodicity is too shaky a basis for the conclusion that a periodic crystal is not capable of spontaneous complication internal structure, and the aperiodic one can draw order from chaos and improve itself. Even if the latter is true, the question remains: why didn’t NaCl turn into an amoeba, and why did the DNA of prokaryotes in a convolution already contain the future man?
N.V. Timofeev-Resovsky:
Darwin did not create the doctrine of evolution, as popularizers often and unreasonably believe. This doctrine was known long before Darwin and was created by Aristotle, Linnaeus, and many others; Darwin had many predecessors in this. The genius of Charles Darwin was that he was the first to see in nature the principle of natural selection, the natural historical mechanism of the evolution of living beings.
If earlier adherents of Darwinism, even denying the “founding father” the discovery of evolution, saw his great merit in the priority of selectogenesis (this priority, as we now know, is also controversial), now the universality of the principle of natural selection is thoroughly questioned. There is “instant” speciation and the matrix mechanism of evolution, and “epidemic” outbreaks of new forms, and the process of “rapid” creation of new forms, which does not correspond to Darwinian evolution, followed by their long-term existence without noticeable changes.
L. S. Berg:
The birth and death of individuals, species, ideas is a catastrophic process. The birth of all these categories is preceded by a long hidden period of development, proceeding on the basis of certain laws, and then immediately comes a leap, saltus, expressed in their birth, spreading on the surface of the Earth and winning their place “under the sun.”
Indeed, if we extend the concept of evolution to the realm of ideas, then we will also see not a simple ascent, but a huge tree with spreading branches, and the throwing out of a new branch-idea is difficult to imagine as a smooth transition - it is always a leap, especially clear for ideas-overshadowings, divine inspirations, news from there...
There is a widespread opinion that speciation is similar to a phase transition, which, like any transition to a new structure or organization, takes a long time to prepare, but occurs spasmodically. The scheme of the evolutionary process according to A. N. Severtsev is qualitatively similar to the phase transition. From a physical point of view, we can also talk about the phenomenon of increasing order in an open system that is far from equilibrium, in which gigantic fluctuations are possible due to the outflow of entropy into the environment.
The latest research (including A. B. Georgievsky and V. Bok) has made it possible to record large mutations that arise immediately as a ready-made adaptation (pre- or super-adaptation). This adaptation may lie dormant if there is no need for it, or it may become an innovation that saves an old species. Life, as if protecting itself from accidents, creates forms that can withstand the most incredible changes in the environment. For example, in the population of a freshwater species there are mutants that, for some unknown reason, are ready to tolerate high salinity. Lowering the temperature of E. coli to almost absolute zero does not kill it: after warming up, its activity is often completely restored. Dried rotifers and tardigrades can sometimes withstand cooling to a temperature of –196 °C. The heat resistance of many organisms also significantly exceeds natural temperatures: there are species that live in thermal springs; the eggs of some crustaceans in a dry state can withstand many hours of heating up to 83 ° C. Ink mold can grow in a 20% solution of copper salts, which usually kills other organisms at concentrations as low as 0.0000001%. One can give a long list of “causeless” super-adaptations that exceed the imaginable needs of the organism and are inexplicable by Darwinian selection.
This suggests the conclusion that selection itself is not among the causes of inertial evolution in the direction of acquiring superadaptations.
The doctrine of evolution is the largest theoretical generalization in modern biology. Biological evolution is understood as an irreversible, progressive and natural process of the historical development of living nature (starting from the moment of the abiogenic emergence of the first living organisms on Earth to the present). As a result of biological evolution, a variety of species of living organisms arose on our planet and the emergence of a biosocial being - man.
Ideas about the natural origin of living beings and their variability already existed in the works of ancient authors. True, the ideas of ancient philosophers were purely speculative, fragmentary and incomplete, often including elements of obvious fantasy.
With the Renaissance, a period of rapid development of biology began, and a huge amount of factual material accumulated. Thanks to numerous geographical discoveries, the lists of animal and plant species known to science are replenished; ideas about the diversity of living organisms are expanding. During this period, biology was armed with comparative and experimental research methods, and microscopy emerged. All this led to the emergence of the principle of gradation, i.e. systems for classifying living organisms according to the degree of their organizational complexity, which was embodied in the construction of “ladders of creatures” or “ladders of natural bodies.” So, C. Bonnet in the middle of the 18th century. proposed his own version of the “ladder”, which begins with “bodies of inanimate nature” - air and water. The following steps are sequentially located: metals, stones, mushrooms, plants, insects, worms, fish, birds, mammals, monkeys and humans. C. Bonnet, like many other authors of similar constructions, did not connect his construction with the idea of the development of nature. We were talking only about the gradation of “natural bodies”.
At the end of the 18th century. In biology and philosophy, transformism is becoming widespread: the idea of the change and transformation of the forms of organisms, the origin of some from others. Transformism played a progressive role in the development of natural science, because opposed creationist ideas. But the views of transformists still lack the idea of evolutionism, i.e. historical (progressive) process of development of living nature.
At the beginning of the 19th century. the first evolutionary theories appear. The J-B theory was of particular importance for the further development of the evolutionary idea. Lamarck (1809), in which, in addition to affirming and substantiating the idea of the historical development of living things, an attempt is made to explain the causes and driving forces of the evolutionary process. Referring the reader to the school course of general biology, where Lamarck's theory is presented in sufficient detail, we note only the fundamentally important ideas expressed by this scientist. Revealing the mechanisms of evolution, Lamarck argued that all living organisms are characterized by variability, which manifests itself as a result of the action of various environmental factors. Along with these rational and productive ideas, Lamarck’s theory contained the irrational idea of “the internal desire of organisms for improvement” and erroneous statements (“changes in organs as a result of their exercise or lack of exercise”, postulating the inheritance of acquired characteristics). However, the theory of J-B. Lamarck was widely recognized by scientists of the 19th century, because it affirmed the principle of the historical development of the organic world and attempted to give a materialistic explanation of the mechanisms of the evolutionary process. The historical role of Lamarck's theory is that it was the first evolutionary theory. It contained ideas that were adopted and developed in the works of other researchers. It is easy to notice the continuity of ideas about the variability of organisms and the role of the environment in the evolutionary process in the series:
J-B theory. Lamarck ® Theory of Charles Darwin ® Synthetic Theory of Evolution.
The largest event in science was the emergence of a scientific theory of the historical development of living nature - the evolutionary theory of Charles Darwin (1859). Darwin's merit lies in the fact that he managed, having determined the driving forces of the evolutionary process, to reveal its essence and build a convincing system of evidence for evolution. This system is the result of a synthesis of enormous factual material not only in biology, but also in other natural and applied sciences, sociology, and demography.
Let us highlight the main provisions of Darwin's evolutionary theory.
1. Artificial selection as a driving factor in the origin and breeding of new varieties of cultivated plants and breeds of domestic animals. The development of this problem played a huge role in the formation of the idea of natural selection, which is the central core of Darwinism.
2. The presence of definite (non-hereditary) and indefinite (hereditary) variability in living organisms. The exceptional importance of hereditary variability in the evolution of living organisms.
3. “The struggle for existence...”, as Charles Darwin himself wrote: “... in a broad and metaphorical sense, including here the dependence of one creature on another, and also implying (more importantly) not only the life of one individual, but also her success in providing herself with offspring.” The struggle for existence arises from the geometric progression of the reproduction of organisms and the numerous obstacles that hinder unlimited reproduction. Darwin emphasized that “the struggle for existence is especially persistent when it occurs between individuals and varieties of the same species.”
4. “Natural selection or survival of the fittest,” as “...the preservation of beneficial individual differences or variations and the elimination of harmful ones.” Darwin believed that "...natural selection acts only by the preservation and accumulation of small hereditary changes, each of which is advantageous to the being preserved."
5. Hereditary variability, the struggle for existence and natural selection are the main driving forces leading to the formation of new species due to “divergence of characters.” Initially, the “divergence of characters” manifests itself in the emergence of varieties within the old species, and as the differences deepen, two (or more) new species arise (the principle of divergence).
6. Natural selection causes the adaptability of species, the appropriateness of the structure and functions of organisms. In other words: evolution is adaptive character. Organic expediency is universal (i.e., fitness is inherent in all living organisms, at all stages of their individual development), but it is relative, because when environmental conditions change, the adaptability of the organism in new conditions will not correspond to them (for example, a delay in snowfall with the onset of winter puts animals in a “difficult situation” that have changed their summer “outfit” to a winter one - the white hare, etc. ). Sharp climatic changes in large areas of our planet often led to the extinction of many species of living organisms (as evidenced by the geological and paleontological records).
7. Progress in the development of the organic world. The essence of progressive evolution lies in the fact that in each historical era, among forms that are well adapted to existing environmental conditions, forms appear that have fundamentally new and more complex types of structure. According to the fossil record, over time, some dominant groups of organisms were replaced (or supplemented) by others that had a higher level of organization than their predecessors (Table 13.3)
Table 13.3
The successive change of dominant forms is based not on particular adaptations, but on adaptations of a broader nature that give organisms general advantages in the struggle for existence and are signs of progress that define groups. The accumulation of such adaptations ensures the progressive course of progressive evolution.
Charles Darwin's theory of evolution, covering the most important problems of biology and giving them a convincing general solution, served as a powerful impetus and laid the foundation for the further development of biology. This theory has undeniable worldview significance. The ideas of evolutionism spread far beyond biology and penetrated into other areas of natural science. Darwinism Presented Today Synthetic Theory of Evolution(STE), which developed by the 40s of the last (twentieth) century.
STE is a direct continuation and development of Darwin's theory. Taking it as a basis, developing the basic Darwinian ideas on the basis of the achievements of modern biological sciences and disciplines (genetics, cytology, ecology, molecular biology, developmental biology, etc.), STE eliminated some of the shortcomings of Darwin’s theory, enriched it with new facts, and supplemented it with some important ideas . The central core of STE, as before (as with Darwin), is the idea of the unity of origin and continuous evolution of living organisms, of the creative role of natural selection.
In 1984, the main provisions of the STE were summarized by N.N. Vorontsov in 11 postulates. Let's consider these postulates.
1. The material for evolution is, as a rule, very small, but discrete changes in heredity - mutations. Mutational variability provides material for natural selection and is random in nature.
2. The leading driving factor of evolution is natural selection, based on the selection of random and small mutations. Hence the name of the selection theory - “selectogenesis”.
3. The smallest evolutionary unit is a population, not an individual (as in Darwin).
4. Evolution is mainly divergent in nature, i.e. one species (taxon) can become the ancestor of several daughter species (taxa), but each species has a single ancestral species, a single ancestral population.
5. Evolution is gradual and long-term. Speciation is thought of as a gradual replacement of one temporary population by a succession of subsequent temporary populations.
6. A species consists of many subordinate morphologically, physiologically and genetically distinct, but not reproductively isolated units - subspecies, populations. This concept is called the broadly polytypic species concept.
7. Gene exchange is possible only within a species. If a mutation has a positive selective value throughout the entire range of a species, then it can spread throughout all its subspecies and populations. This leads to a brief definition of a species as a genetically integral and closed system.
8. Since the criterion of a species is its reproductive isolation, it is natural that this criterion is not applicable to forms without a sexual process, for example, to agamic and parthenogenetic organisms. Thus, STE left out of species status a huge number of prokaryotes that do not have the sexual process, as well as some specialized forms of higher eukaryotes that have lost the sexual process for the second time.
9. Macroevolution, or evolution at a level above the species, proceeds only through microevolution. According to STE, there are no patterns of macroevolution that are different from microevolutionary ones, although there are phenomena (parallelism, convergence, analogy, homology) that are easier to study at the macroevolutionary level.
10. Each systematic unit: species, genus, etc., must have a single root, monophyletic origin. This is a prerequisite for the very right to exist of the group in question. After all, evolutionary systematics builds its classification based on their kinship. And according to the fourth postulate of STE, only those groups that come from the same evolutionary branch are related. If a taxon suddenly has two different branches in its ancestors, it should be divided.
11. Based on all the postulates mentioned, it is clear that evolution is unpredictable: it is not directed towards some final goal, i.e. is non-finalistic in nature.

In 2018, the Nativity Fast will begin on November 28. During this period, Orthodox believers prepare to celebrate Christmas...

Starting a family is the dream of most women. They want to have a loving husband and a bunch of kids. But it's not always a relationship...

This article contains: the most powerful prayer for divorce - information taken from all over the world, the electronic network and...

Information site about icons, prayers, Orthodox traditions. Prayer for scandals and quarrels in the family, with husband, with children...

What would New Year be without champagne, tangerines, Olivier, aspic and everyone’s favorite “Herring under a fur coat”. With the last one...

Let's prepare the necessary ingredients for the cookies. The first thing to do is put the water to boil. We need...

Is it possible to register an employee for the position of financial director - chief accountant? The chief accountant claims...

The head of a small business can easily manage the budget independently. CHECKED! If you manage...

The creation of new projects involves a preliminary economic justification for their feasibility, subsequent...
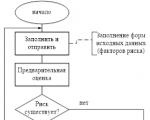
Reporting is generated by the RM, is agreed upon (approved) by the Risk Committee under the Management Board and transmitted to...

At the edge of a large, very large meadow, on a long emerald blade of grass lived a tiny Ladybug. God's little...

Nowadays, it is quite common for people to turn to the stars. With the help of a horoscope a person can find out...

Business or friendly. If you were pursuing a stranger, it means your level of trust in the female sex...

according to Freud's dream book If you dreamed about how you were fishing, it means that in real life you can hardly switch off...

Starting a family is the dream of most women. They want to have a loving husband and a bunch of kids. But it's not always...

This article contains: the most powerful prayer for divorce - information taken from all over the world, electronic...