Філософське бачення проблеми
Сенс життя пов'язаний із питанням «Заради чого жити», а не питанням про те, як підтримувати життя. Ставлення людини як...
До написання статті мене підштовхнули безперервні розмови про те, що українці - це слов'яни, а росіяни - це взагалі не слов'яни, а давно вже монголи.
Природно, ініціаторами подібних суперечок виступають так звані українські патріоти. У цьому висновки робляться з урахуванням теорій деяких новоявлених істориків, досі невідомих історичних документів тощо. Але крім історії, а найчастіше псевдоісторії, є ще й така наука, як генетика, а з генетикою дорогі мої не посперечаєшся. Так що хочемо ми того чи ні, але генотип у нас один.
Що таке гаплогрупа?
Y-хромосомні гаплогрупи, що стали популярними у колах, стурбованих біополітикою, – це статистичні маркери, що дозволяють зрозуміти походження людських популяцій. Але в більшості випадків такий маркер нічого не говорить про етнічну чи расову приналежність окремої людини (на відміну від інших методик аналізу ДНК). Бачити в сукупності носіїв конкретної гаплогрупи етнос, субетнос, расу чи іншу єдність подібного роду, намагатися сколотити якусь ідентичність на цьому ґрунті – це безглуздя. І, зрозуміло, гаплогрупа аж ніяк не «відбивається на кшталт людини».
Особливість Y-хромосоми в тому, що вона передається від батька до сина практично в незмінному вигляді і не відчуває «змішення» та «розведення» материнської спадковістю. Це дозволяє використовувати її як математично точний інструмент для визначення походження батьківської лінії. Якщо термін «династія» має якийсь біологічний сенс, це саме успадкування Y-хромосоми. (За посиланням можна знайти детальне, але просте розуміння пояснення феномена)
Інша справа Y-хромосома: вона складається з генів, які безпосередньо відповідають за чоловічу систему відтворення, і найменший дефект, як правило, робить чоловіка стерильним. "Шлюб" далі не передається, і Y-хромосома "самоочищається" в кожному поколінні.
Але крім шкідливих мутацій, у чоловічій хромосомі іноді трапляються нейтральні, ігноровані природним добором. Вони зосереджені у «сміттєвих» ділянках хромосоми, які є генами. Деякі з цих мутацій, що сталися від 50 до 10 тисяч років тому, виявилися зручними маркерами для позначення стародавніх популяцій предкових, які згодом розселилися по всій Землі і сформували сучасне людство.
Y-хромосомна гаплогрупа визначає сукупність чоловіків, що об'єднані наявністю такого маркера, тобто. що походять від загального предка-патріарха, у якого багато тисяч років тому трапилася специфічна мутація Y-хромосоми.
http://en.wikipedia.org/wiki/Human_Y-chromosome_DNA_haplogroup
ПОХОДЖЕННЯ ГАПЛОГРУПИ R1a1 - ПІВДЕНЬ РОСІЇ!
Будь-який сучасний етнос складається з представників кількох як мінімум двох-трьох Y-хромосомних гаплогруп.
http://en.wikipedia.org/wiki/Y-DNA_haplogroups_by_ethnic_group
Географічне поширення гаплогруп пов'язані з історією міграцій давніх популяцій, які стали предковими для етносів чи груп етносів. Наприклад, гаплогрупу N3 можна назвати «фінно-угорською»: якщо вона зустрічається у представників якоїсь місцевості, то значить, у минулому населення тут поєднувалося з фінно-уграми. Чи сюди прийшли вже «змішані» племена.
Вивчення статистики гаплогруп дозволило антропологам відновити картину міграцій людських популяцій останні десятки тисяч років, починаючи з африканської прабатьківщини. Але ці дані можна використовувати для викриття різноманітних расистських і ксенофобських міфів.
Етногеографічне поширення гаплогрупи R1a
В даний час високі частоти гаплогрупи R1a знаходяться в Польщі (56% населення), Україні (від 50 до 65%), європейській частині Росії (від 45 до 65%), Білорусі (45%), Словаччині (40%), Латвії. (40%), Литві (38%), Чехії (34%), Угорщини (32%), Хорватії (29%), Норвегії (28%), Австрії (26%), Швеції (24%), північний схід Німеччини (23%) та Румунії (22%).
Найбільше поширення має Східна Європа: серед лужичан (63 %), поляків (бл. 56 %), українців (бл. 54 %), білорусів (52 %), росіян (48 %), татар 34 %, башкир (26 %) ) (у башкир Саратовської та Самарської обл. до 48%); і в Центральній Азії: у худжандських таджиків (64%), киргизів (63%), ішкаші (68%).
Галогруппа R1a найбільш характерна для слов'ян. Наприклад, у росіян поширені такі гаплогрупи:
R1a – 51% (слов'яни – арії, поляки, росіяни, білоруси, українці)
N3 – 22% (фінно-угри, фіни, прибалти)
I1b – 12% (нормани – германці)
R1b - 7% (кельти та італіки)
11a - 5% (теж скандинави)
E3b1 - 3% (середземноморці)
Найпоширеніший гаплогрупи в українців:
R1a1 – близько 54 % (слов'яни – арії, поляки, росіяни, білоруси, українці)
I2a – 16,1% (балканські народи, фраційці, іллірійці, румуни, албанці, греки)
N3 - 7% (фінно-угри)
E1b1b1 - 6% (африканські народи, єгиптяни, бербери, кушніри)
N1c1 - 6% (сибірські народи, якути, буряти, чукчі)
Як показують дослідження, за маркерами Y-хромосоми протестовані українці найбільш генетично близькі до південно-західних російських, білорусів і східних поляків, що сусідять з ними. Три слов'яномовні народи (українці, поляки та росіяни) формує окремий кластер по Y гаплогрупах, що вказує на загальне походження перелічених етносів.
Міфи.
Всім відомий міф про те, що росіяни – це значною мірою нащадки монголів, які поневолювали Русь у давнину. Статистика гаплогруп не залишає від цього міфу каменю на камені, оскільки типові «монголоїдні» гаплогрупи C і Q у росіян взагалі не зустрічаються. Значить, якщо монгольські воїни і приходили колись на Русь з набігами, то всіх спійманих жінок вбивали або забирали з собою (як кримські татари в більш пізні часи).
Ще один поширений міф - про те, що росіяни в Центральній та Північній Росії - це здебільшого не слов'яни, а нащадки фінно-угорських аборигенів, у морі яких, нібито, розчинилися нечисленні слов'яни. Звідси виводять «російське пияцтво», «російську лінь» тощо. Тим часом, частка «фінської» гаплогрупи N3 у росіян у Центральній Росії становить приблизно 16% (у малонаселених регіонах на північ від Москви подекуди доходить до 35%, а в густонаселених регіонах на південь і на захід від Рязані - скорочується до 10%). Тобто. із кожних шести батьків лише один був фіном. Можна припустити, що і за материнським генофондом співвідношення приблизно таке саме, оскільки слов'яни та фінно-угри, як правило, мирно співіснували.
До речі, у фінів Фінляндії гаплогрупа N3 представлена приблизно у 60% населення. Отже, з кожних п'яти батьків двоє були не «споконвічними фінами», а «проїжджими молодцями», можливо – збирачами ясаку з Новгорода. У етнічних естонців та латишів «частка фінських батьків» ще менша – приблизно 40%. "Проїжджі молодці" німецького та слов'янського походження явно домінували над гарячими естонськими хлопцями. Проте ті полюбилися литовським дівчатам: литовці, незважаючи на індоєвропейську мову, є нащадками фінно-угрів на ті ж 40%.

У етнічних українців «частка фінських батьків» теж є, хоча й утричі менше, ніж у росіян. Однак фінно-угорські племена в Україні не жили, і ця частка занесена із Центральної Росії. Але якщо «частка фінської крові» в етнічних українців лише втричі менша, ніж у росіян, то, отже, як мінімум третина з них – це нащадки російських батьків. Мабуть, у минулому «несвідомі» південноруські дівчата любили повеселити з «москальськими окупантами». У той час як українські хлопці розважалися в Запорізькій Січі в суто чоловічій компанії, їхні сестри та дочки знаходили розуміння у привітних суворовських чудо-богатир із важкими фінськими Y-хромосомами.
Допомагаючи зрозуміти неспроможність тих чи інших міфів, гаплогрупи, у свою чергу, здатні породити нову міфотворчість. Знаходяться люди, які надають їм расового змісту. Важливо зрозуміти, що самі по собі гаплогрупи не можуть бути критерієм расової, етнічної чи субетнічної ідентичності. Стосовно конкретної людини вони взагалі ні про що не говорять. Наприклад, жодної адекватної спільності, що об'єднує людей із «арійської» гаплогрупи R1a1, сформувати не вдасться. І навпаки, жодної об'єктивної різниці між російськими, що живуть в одному регіоні, носіями «фінської» гаплогрупи N, і російськими, носіями «арійської» гаплогрупи R1a, не існує. Весь решта генофонду у нащадків «предкових чоловіків-фінів» та «предкових чоловіків-аріїв» давно перемішався.
З більш ніж 20 000 генів людського геному, в Y-хромосому входить всього близько 100 штук. Вони кодують в основному будову та функціонування чоловічих статевих органів. Жодної іншої інформації там немає. Риси обличчя, колір шкіри, особливості психіки та мислення прописані в інших хромосомах, які при наслідуванні проходять через рекомбінацію (батьківські та материнські ділянки хромосом перемішуються випадковим чином).
Якщо представники якогось етносу ставляться до кількох гаплогруп, це не означає, що цей етнос є механічне з'єднання популяцій з різним генофондом. Весь решта генофонду, крім Y-хромосом, у них буде перемішаний. Тонкі відмінності між представниками різних російських гаплогруп можуть бути цікаві лише людям, які професійно спеціалізуються на мінеті.
І навпаки, люди з однієї гаплогрупи можуть належати до різних етносів і навіть різних рас, мати принципові відмінності з точки зору генотипу і фенотипу.
Наприклад, рекордсменами за присутністю «арійської» гаплогрупи є такі несхожі один на одного народи, як поляки (56,4%) та киргизи (63,5%). «Арійська» гаплогрупа зустрічається більш ніж у 12% євреїв-ашкеназі, причому не в якихось «напівкровок», а у справжнісіньких, типових представників свого етносу.
Якщо російські моряки, відвідавши Анголу, «подарують» тубільці дитини-хлопчика, то в нього, і у всіх його нащадків по чоловічій лінії, буде гаплогрупа батька. Зміниться 1000 поколінь, нащадки за всіма параметрами перетворяться на типові ангольці, але все одно носитимуть «арійську» Y-хромосому. І жодним іншим чином, крім аналізу ДНК, цей факт виявити не вдасться.
У далекому минулому носії гаплогрупи R1a1, предки сучасних індоєвропейців, вирушили з Південної Росії та Пріуралля на освоєння Європи, Близького Сходу, Ірану, Афганістану, Індії та інших суміжних країн, населенню яких вони нав'язали свої звичаї та передали свою мову. Але якщо їхня історична успішність була якось пов'язана з просунутою біологією (припустимо), то коренилася вона не в особливостях Y-хромосоми, а в інших генах, які були у предкової популяції. З певною гаплогрупою цей "просунутий генофонд" був пов'язаний лише статистично. У сучасних представників гаплогрупи R1a1 ці «просунуті» гени можуть бути відсутніми. Володіння «арійської» хромосомою не відбивається «на кшталт».
Ті ділянки Y-хромосоми, які є маркерами виділення гаплогруп, власними силами нічого не кодують і жодного біологічного сенсу немає. Це маркери у чистому вигляді. Їх можна порівняти з помаранчевим та зеленим світлодіодами у фільмі «Кін-дза-дза», які використовувалися для ідентифікації чатлан та пацаків, причому жодної іншої різниці, крім кольору лампочки, між цими «расами» не було. Отже наявність «арійської» гаплогрупи сама собою не гарантує людині як арійських мізків, і навіть арійського члена («расисти-минетчики» можуть бути розчарованими).
У ДНК людини 46 хромосом, кожна людина успадковує половину від батька хромосом, половину - від матері. З 23 хромосом, отриманих від батька, тільки в одній – чоловічій Y-хромосомі – міститься набір нуклеотидів, який передається з покоління до покоління без змін протягом тисячоліть.
Генетики називають набір ДНК, що передається у спадок без змін - гаплогрупою.
ДНК дослідження об'єднали всіх людей Землі в генеалогічні групи і позначили їх літерами. Люди однієї гаплогрупи мають одного спільного їм предка у далекій передісторії.
Гаплогрупа, з її спадкової незмінності, однакова в усіх чоловіків одного народу.
У кожного біологічно самобутнього народу є своя гаплогрупа, що відрізняється від гаплогруп інших народів. Гаплогрупа – це є генетичний маркер всього народу.


Ось список Y-ДНК гаплогруп ВСІХ типів,які виникли на певних територіях та через деякий час розійшлися територією Європи та Азії.
Гаплотип:
До— виник 40 000 років тому в північному Ірані.
Т- виник 30000 років тому вздовж берегових ліній Червоного моря та/або навколо Перської затоки.
J- З'явився 30000 років тому на Близькому Сході.
R- виник 28000 років тому у Центральній Азії.
E1b1b- виник 26000 років тому у Північно-Східній Африці.
I -виник 25 000 років тому на Балканах.
J1 -виник 20000 років тому в горах Таврос/Загрос.
J2- З'явився 19000 років тому в північній Месопотамії.
E-M78- виник 18000 років тому у північно-східній частині Африки.
R1b- виник 18000 років тому вздовж берегової лінії Каспійського морята/або Центральної Азії.
R1a- виник 17000 років тому на півдні Росії.
G- З'явився 17000 років тому на Близькому Сході.
I2- З'явився 17000 років тому на Балканах.
E-V13- виник 15000 років тому у південному Лівані чи Північній Африці.
I2b -виник 13 000 років тому в Центральній Європі.
N1c1 -виник 12000 років тому в Сибіру.
E-M81- виник 11000 років тому у Північно-Західній Африці.
I2a- З'явився 11000 років тому на Балканах.
G2a— виник 11000 років тому у Лівані чи Анатолії.
R1b1b2- виник 10000 років тому на півночі чи півдні Кавказу.
I2b1- З'явився 9000 років тому в Німеччині.
I2a1- виник 8000 років тому у Південно-Західній Європі.
I2a2- виник 7500 років тому у Південно-Східній Європі.
I1- З'явився 5000 років тому в Скандинавії.
R1b-L21- виник 4000 років тому у Центральній та Східній Європі.
R1b-S28- виник 3500 років тому у районі Альп.
R1b-S21- виник 3000 років тому у Фрісландії (Фризія) на узбережжі Північного моря та у Центральній Європі.
I2b1a- виник менше 3000 років тому у Великій Британії.
Аналіз генофонду Європи розпочнемо з розгляду геногеографії окремих гаплогруп. Почасти це є продовженням попереднього розділу, в якому коротко підсумовувалося те, що було відомо про поширення гаплогруп Y-хромосоми до нашого дослідження, але тепер ми розглянемо сучасні, докладні карти розповсюдження всіх основних європейських гаплогруп та тих їх субваріантів, для яких вже відомі частоти низці популяцій Європи.
У нашій базі даних Y-base міститься майже вся опублікована інформація про частоти гаплогруп Y-хромосоми у світі (і в Європі зокрема), але далеко не вся ця інформація придатна для вивчення географічного розповсюдження гаплогруп. Як і при створенні інших геногеографічних атласів, для створення карт поширення Y-хромосоми в Європі дані були ретельно відібрані та створені масив даних для карт Y-хромосоми у Європі. Розглянемо, за якими критеріями цей масив даних формувався.
МАСИВ ДАНИХ ДЛЯ КАРТ: ПОПУЛЯЦІЇ.
Для надійного визначення частоти однобатьківських генетичних маркерів (Y-хромосоми та мтДНК) потрібен обсяг вибірки вдвічі більший, ніж для аутосомних маркерів – тому нижня межа вибірки має бути не менше 70-100 зразків. Маленькі вибірки або поєднувалися з географічно сусідніми вибірками того ж народу, або (якщо об'єднання було неможливо) не включалися до аналізу. В результаті середній обсяг вибірки масиву даних для карт становить N=136 зразків. Але робилися й винятки із правила. За деякими народами чи регіональними групами в межах народу, які для повноти картини хотілося обов'язково представити на карті, були доступні лише невеликі вибірки. Тоді, скріпивши серце, доводилося включати до масиву для карт та вибірки меншого розміру. Але такі винятки робилися рідко - з 251 картографованих популяцій лише десята частина (29 популяцій) мають обсяг вибірки нижче за N=50, а переважна більшість (192 вибірки) мають обсяг N=70 і вище.
Якщо якогось народу були дані щодо його регіональним популяціям, то картографування не використовувалися вибірки цього народу, котрим географічна прив'язка невідома (якщо автори при публікації вказали лише приналежність народу).
У результаті масив для карт включив дані по 251 населенням (вибіркам), і сумарне число зразків у всіх вибірках становило 34 294 зразків. Варто зазначити, що значна частина цих даних отримана завдяки дослідженням нашого колективу під керівництвом автора цих рядків: 45 популяцій та 4880 зразків. Це наші дані щодо російських популяцій, українських, білоруських та деяких додаткових російських популяцій, популяцій Північного Кавказу, а також наші ще неопубліковані дані щодо популяцій кримських і казанських татар, мокші та ерзі. (Дані за частотами гаплогруп з трьох опублікованих статей наведені в таблицях 2.1, 2.2, 2.3.) Виходить, що з усього масиву даних мінливості Y-хромосоми в Європі кожна п'ята популяція і кожен сьомий зразок вивчені нашим колективом (рис. 2). популяції показані синіми гуртками), хоча дослідженнями в цій галузі займалося і займається кілька десятків лабораторій по всій Європі. Якщо додати, що Європа є найбільш вивченим регіоном світу, то це частково пояснює, чому саме нашим колективом зроблено узагальнюючий аналіз генофонду Європи. Для побудови карт використані також дані Y-base не тільки по європейських популяціях, але і по популяціях прилеглих регіонів, що входять до ареалу, що картографується.
Крім 251 популяцій Європи, вивчених за панелями гаплогруп, що охоплюють весь їх спектр, ще близько 200 популяцій було вивчено за субваріантами лише однієї гаплогрупи. В основному це статті, присвячені гаплогруп R1b і R1a, і ряд статей з гаплогруп N, E, і J. Як правило, вибірки, використані в цих статтях для поглибленого генотипування окремих гаплогруп, раніше вже вивчалися по всьому спектру гаплогруп. Рис. 2.1. дає уявлення про щільність покриття Європи та суміжних регіонів популяціями, вивченими не за однією гаплогрупою, а по всьому їх спектру зі звичайним рівнем філогенетичного дозволу.
МАСИВ ДАНИХ ДЛЯ КАРТ: ГАПЛОГРУПИ.
Це поняття «звичайного філогенетичного рівня» вивченості гаплогруп дуже умовно, і цей рівень зростає з кожним роком. Якщо в 2000 році число гаплогруп у звичайній статті не дотягувало до десятка, то в 2010-ті роки більшість статей використовують панелі з 40-70 гаплогруп. Звичайно, починаючи приблизно з 2013 року кількість відомих гаплогруп, виявлених завдяки повному секвенуванню, обчислюється вже тисячами. Але виявлення їх - лише перший крок, який вимагає безліч наступних кроків і зусиль з аналізу великого масиву різних популяцій на наявність нових гаплогруп. І доки не проведено визначення їх частот у широкому спектрі популяцій, значення цих нововідкритих гаплогруп для досліджень генофонду невелике.
Об'єднуючи в одній таблиці дані з десятків статей різних років, неминуче виявляється, що деякі гаплогрупи вивчені в дуже великій кількості популяцій, а деякі лише для небагатьох популяцій. На щастя, для картографування це не є великою проблемою (уже в двох-трьох десятках популяцій вивчені майже всі відібрані для картографування гаплогрупи, а такого числа опорних точок зазвичай достатньо для виявлення основних трендів). Але все ж таки при розгляді карток варто мати на увазі, що подробиця вихідних даних для різних гаплогруп неоднакова.
Для картографування відбиралися гаплогрупи, що становлять помітну частину генофонду хоча б у деяких популяціях Європи та при цьому вивчені, як щойно було сказано, у кількох десятках популяцій. Цим критеріям задовольнили 40 гаплогруп: E1b1a1-DYS271, E1b1b1a1b-L142.1, E1b1b1a1c-V22, E1b1b1b1-M81, E1b1b1c-M123, G2a1-P16 L118, I2a-P37.2, I2b-L35, J1-L255, J1c3-P58, J2-L228, J2a-L152, J2a3b-M67, J2a3h-L207.1, J2b-L282, L-M11, N1b-P43, N1c-M42, O3 Q-M242, R1a1-L120, R1a1a1g-M458, R1a-M558, R1a-Z284, R1a-Z93, R1b1a1-M478, R1b1a2-L265, R1b-L10, R1b1a2a1a1 1b1a2a1a1b-P312, R1b1a2a1a1b2a- M153, R1b1a2a1a1b2b1-M167, R1b1a2a1a1b4b-M222, R1b1c-V88, R2a-L266, T-L206.
Таблиця 2.1. Наші дані про частоти гаплогруп Y-хромосоми в російських популяціях
Таблиця 2.2. Наші дані про частоти гаплогруп Y-хромосоми в популяціях балто-слов'янських народів (опубліковані в ).

Таблиця 2.3. Наші дані про частоти гаплогруп Y-хромосоми у популяціях Кавказу


Мал. 2.1. Географічне положення популяцій Європи та суміжних регіонів, вивчених за поліморфізмом Y-хромосоми. Синіми кружальцями показані наші власні дані, зеленими – дані з літератури.
ГАПЛОГРУПАR1 a(без підрозділу на субгалузі).
Як зазначалося в огляді попередніх робіт, гаплогрупа R1 aє однією з двох найчастіших гаплогруп Європи. Більше того, вона є і найпоширенішою гаплогрупою в Євразії в цілому. Для її генотипування використовуються різні маркери - M198, M17, SRY1542, філогенетична різниця між якими майже відсутня, і відомо безліч інших маркерів, що визначають цю гаплогрупу, тому для її позначення може використовуватися будь-який з них. У масиві даних для карток вона фігурує як R1 a- L120 .

На рис. 2.2 можна побачити чіткий тренд географічного поширення R1 a- L120 : хоча вона поширена по всій Європі, але велика зона максимальних частот знаходиться в Східній Європі, причому в цих популяціях вона становить майже половину генофонду. На сході зону максимуму обмежує Волга, причому по всьому її протязі – від Верхньої Волги до низовин. На півдні зона максимуму сягає майже Чорного моря, на північному заході – до Балтійського моря, а на південному заході обмежується Карпатами та Альпами. Але найбільш цікавими є межі цієї області на заході: хоча ніяких географічних бар'єрів там немає, зона високих частот R1a все одно досить чітко обмежена, і падіння частоти на якихось двохстах-трьохстах кілометрах досягає майже 30% (від приблизно 35-45% у поляків та сорбів до 10-15% у німців). Аналізу цього «польсько-німецького генетичного кордону» було присвячено кілька статей.
Я не згадав ще межу зони максимальних частот R1a на півночі - хоча перепад частот там не такий виражений, як на заході (не від червоних до зелених, а тільки від червоних до жовтих тонів, тобто відсотків на п'ятнадцять), але північний кордон теж виявляється на карті цілком рельєфно. У разі вона проходить всередині ареалу одного народу – російського – маркуючи відмінності генофонду Російської Півночі від інших російських популяцій. Ці жовті тони помірних частот займають як Російська Північ, а й усе простір Східної Європи між Волгою і Уралом, і лише за Уральським хребтом частоти R1 aпадають до так само низьких значень (зелені тони на карті), що й у Західній Європі, Скандинавії, Балканах та Передній Азії.
СУБВАРІАНТИ ГАПЛОГРУПИR1 a
У межах такого величезного ареалу гаплогрупи R1a-L120 Безсумнівно, існує майже так само величезна кількість субваріантів цієї гаплогрупи, причому багато хто з них може мати чіткі і порівняно невеликі географічні ареали. Але протягом тривалого часу субтипи в межах R1 aбули невідомі. Поки що завдяки роботам лабораторії Peter Underhill у Стенфордському університеті не було відкрито спочатку перші два, а потім ще кілька субваріантів. Звичайно, нині кількість відомих субваріантів R1 aобчислюється сотнями: кожен зразок, для якого проведено повне секвенування всього геному або Y-хромосоми, утворює свою гілку на філогенетичному дереві. Але поки хоча б для деяких з цих варіантів не проведено скринінгу їх поширеності в популяціях і не визначено частоти їхньої зустрічальності в різних популяціях Європи, побудова геногеографічних карт неможлива. Зараз про ареали поширення цієї субгілок можна міркувати лише вороже, грунтуючись на часом хиткі відомості про місця походження тих одиничних зразків, за якими кожна з цих сотень гілок була відкрита. Інтерес до цієї гаплогрупи настільки великий, що, звичайно, такий популяційний скринінг буде проведено різними лабораторіями світу (у тому числі і нашим колективом) у найближчі роки, і тоді геногеографія субваріантів R1 aстане на порядок детальніше. Але на даний момент ми обмежені тими маркерами, за якими проведено масовий популяційний скринінг у статтях та роботах нашого колективу: це три основні маркери європейської гілки. R1 aі маркер азіатських гілки.
АЗІАТСЬКА ГІЛКАR1 A- Z93.
Слід зазначити, що розподіл на європейську та азіатську гілки проявляється на філогенетичному дереві. R1 aдуже чітко. Маркером азіатської гілки є Z93 (та її синоніми), й у Європі ця гілка майже відсутня. На підтвердження цього наведемо карту її поширення (рис. 2.3.) на якій видно лише низькі частоти (1-3%) і лише в небагатьох популяціях Східної Європи, і перейдемо до розгляду європейських субваріантів.

Мал. 2.3. Географія гаплогрупыR- Z93 Y-хромосоми вЄвропі.
ЄВРОПЕЙСЬКА ГІЛКАR1 A-М458.
Перший, R-М458, виділяє центрально-європейський субваріант у загальному масиві гаплогрупи R1aзаймає половину Європи. Тому після відкриття цього маркера, ми спеціально генотипували його у багатьох слов'янських та північно-кавказьких популяціях, що дозволило вперше побудувати докладну карту його поширення з урахуванням Східної Європи та Кавказу (рис. 2.4).

Карта демонструє найвищі частоти цього маркера у західних слов'ян (поляків та чехів), де він становить чверть генофонду, і дещо знижені частоти в навколишніх популяціях. Поширення цього маркера на захід обмежено прилеглими популяціями німців (ймовірно, західнослов'янські групи, що асимілювали, раніше населяли ці території). Натомість на схід зони високих частот гаплогрупи R-M458поширюється далеко, захоплюючи весь ареал східних слов'ян. Характерно, що ця східна експансія більш виражена у лісостеповій зоні (в українців), ніж у лісовій (білоруси та росіяни). Ще цікавішим є обмеженість експансії на південь: у південних слов'ян і неслов'янських народів Балкан (угорці, румуни) частота цієї гаплогрупи мала, причому простежується відповідність генетичної «кордону» високих частот R-M458та географічного кордону Карпат. На північ від цього гірського хребта розташована зона максимальних частот гаплогрупи, а на південь від Карпат частота різко знижується. Очевидно, Карпати виступили у ролі географічного бар'єру для потоку генів, перешкоджаючи поширенню цієї гаплогрупи на Балкани. Зазначимо, що на відміну від R1 aв цілому, що зустрічається хоча б з низькими частотами у будь-якому регіоні Європи, R- M458 поширена тільки по всій Східній Європі, сусідніх територіях Західної Європи та на Балканах, але практично повністю відсутня на більшій частині Західної Європи – Піренейському півострові, Франції, Британських островах, значній частині Апенинського та Скандинавського півостровів, а також відсутня за межами Європи – і на Передній Азії і в Сибіру.
ЄВРОПЕЙСЬКА ГІЛКАR1 A-М558.
Другий європейський субваріант R1 a позначається за назвою маркера R- M558 (Мал. 2.5.) Філогенетично він братський (для мтДНК у таких випадках прийнято говорити «сестринський», але тут йдеться про Y-хромосому) для R- M458 . Обидва ці маркери поширені подібним чином: R- M558 теж поширений переважно у Східній Європі, теж рідше зустрічається на Балканах, заходить із низькими частотами у сусідні області Західної Європи, і так само, як і R- M458 , практично відсутня за межами Європи Ареали цих субтипів перекриваються і в зоні їх максимальних частот у західних і східних слов'ян. Основна відмінність між патернами поширення R- M458 і R- M558 полягає лише у більшій приуроченості до західної чи східної частини їхнього загального ареалу.

R- M458 має схильність на захід ареалу. У західних слов'ян зустрічається в середньому із частотою 25% (тобто чверть генофонду), а у східних слов'ян у середньому зустрічається із частотою 15%, хоча й підвищується до 23% у західних білорусів та в деяких українських популяцій (але не західних, а центральних).
R- M558 має схильність на схід їхнього загального ареалу. Він зустрічається з частотою понад 30% в обох вивчених за цим маркером популяціях південних росіян; становить 20-25% у білорусів, поляків, словаків, західних українців; знижується нижче (або значно нижче) 20% у центральних росіян, чехів, центральних та східних українців та чехів. Втім, R- M558 вивчений у значно меншій кількості популяцій, ніж R- M458, і тому карта його поширення може значно уточнитися у майбутньому. Ми бачимо, що навіть ця основна відмінність між патернами поширення R- M458 і R- M458 виявляється у невеликих відмінностях частот і має досить складну географію.
Цей приклад ясно показує, що підвищення філогенетичного дозволу далеко не завжди призводить до підвищення географічного дозволу: хоча в межах європейської галузі R1 aі вдалося виділити два субтипи, та їх географічне поширення дуже схоже. Очевидно, вони з'явилися у родинних популяціях і поширювалися разом, під час тих самих міграцій. Або, виникнувши навіть у різних частинах ареалу популяцій, інтенсивно обмінюваних міграціями друг з одним, вони поширилися не більше цього ареалу. Звичайно, підвищувати філогенетичний дозвіл необхідно, і, як правило, субтипи, що виділяються, мають більш чітку географічну прив'язку, ніж гілка в цілому; субтипи наступного рівня мають ще вужчу зону поширення в межах зон субтипів попереднього рівня і так далі. Але ця географічна чіткість залежить не тільки і навіть не так від рівня філогенетичного дозволу, як від структури генофонду, від історично заданого співвідношення ізоляції та метисації, тобто, в термінах популяційної генетики, від балансу дрейфу генів та міграцій.
Якщо цей баланс зрушений у бік ізоляції (як у Кавказі чи Сибіру), то частоти гаплогруп різко відрізнятимуться між популяціями, гаплогрупи матимуть географічно вузькі ареали, мало перекриваються друг з одним, і гаплогрупи досягатимуть дуже високих частот у «своїх» популяціях . Якщо ж баланс зрушений у бік міграцій, то частоти будуть відрізнятися не настільки драматично, ареали будуть досить широкими і перекриваються один з одним, а субгаплогрупи досягати не надто високих частот, тому що решта генофонду буде представлена іншими гаплогрупами, ареали яких заходять на цю ж територію. Всі ці риси добре видно на прикладі R- M458 і R- M558 тому, що зсув балансу у бік не ізоляції, а міграцій особливо характерний саме для Європи.
ЄВРОПЕЙСЬКА ГІЛКАR1 A-Z284.
Натомість ще один субваріант R1 a– гаплогрупа R- Z284 (рис. 2.6) має зовсім інший патерн, який можна назвати «скандинавським» або «північноєвропейським». За наявними на даний момент даними його частота становить 20% у Норвегії, 7% у Данії, і по 3% в Англії та Швеції, а за межами Північної Європи R- Z284 зустрінута лише у поодиноких зразків. Ці частоти напевно скоригуються під час вивчення нових популяцій та збільшення обсягів вибірок вже вивчених народів, але загальна приуроченість до популяцій Північної Європи безсумнівна вже сьогодні.

ГАПЛОГРУПАR1 b(В ЦІЛОМУ)
Гаплогрупа R1 b, «братня» по відношенню до гаплогрупи R1 aі друга серед найчастіших у Європі, має також широкий ареал (Рис. 2.8), проте основна зона її максимальних частот посідає Західну Європу. Важливим внеском у вивчення генофонду Європи стало відкриття нових інформативних SNP маркерів у межах однієї із гілок гаплогрупи R1 b- Гаплогрупи R-М269(на картах позначена за допомогою філогенетично близького маркера як R-L265 ). Це масштабне дослідження виконано великим міжнародним колективом за участю автора та опубліковано у .
R1 bв цілому ( R- L10 , Мал. 2.8) яскраво показує, що це основний компонент, що становить більше половини Y-хромосомного генофонду Західної Європи. Лише населення південної частини Апенінського півострова несе знижені частоти цієї гаплогрупи, оскільки в них переважають середземноморські гаплогрупи, та ще й Скандинавія з географії гаплогруп відноситься не до Західної Європи, а формує самостійний північний домен, в якому переважає інша гаплогрупа. I1 .
Гаплогрупа R1 bнавіть більшою мірою властива Західній Європі, ніж R1 aвластива Східній, оскільки R1 bу багатьох популяціях (Британські острови, баски та інших популяцій Піренейського півострова) становить навіть половину, а більше двох третин генофонду. В цілому, тренд гаплогрупи R1 b- максимальні частоти у басків і поступове зниження частоти на схід від Піренейського півострова - дуже нагадує географію класичного маркера Rh-d(резус фактор). Сплеск частоти Сході Європи відбиває підвищену частоту R1bу деяких популяцій башкир, відому із роботи [Лобов, 2009]. Втім, нашим колективом зараз завершується докладне дослідження різноманітності Y-хромосоми у популяціях башкир (понад 1000 зразків). У цьому дослідженні виявлено, що частота R1 bвелика лише у небагатьох родових групах, тоді як більшість родових груп башкир характерні інші гаплогрупы, а частоти R1 bу них невеликі. Тому сплеск R1 bСході Європи відбиває не загальноєвропейську закономірність, лише локальні закономірності походження окремих родових груп степових популяцій Євразії.
Хоча R1 bдосягає високих частот в основному в Західній Європі, карта (рис. 2.8.) показує, що вона (як і R1 a) поширена повсюдно, займаючи весь картографований ареал і йдучи далеко за межі Євразії. Вважається що R1 bмає передньоазіатське походження, і різні її гілки поширилися різними куточками Старого Світу, але особливо «пощастило» лише тій гілки, яка поширилася в Європі і досягла в ній таких значних частот.

На малюнку показано лише гілки, які докладно розглядаються за допомогою геногеографічних карт; структура схеми відповідає повному дереву гаплогрупи R1b-L10 за версією ISOGG на жовтень 2015 року.

СУБВАРІАНТИ ГАПЛОГРУПИR1 b
Мапи на рис. 2.9, 2.10 та 2.11 показують три основні гілки R1 b(Рис. 2.7.), які – з безліччю застережень – можна назвати африканською, степовою євразійською та західноєвропейською гаплогрупами. Дійсно, коли ми дивимося на карту R1 bзагалом (рис. 2.8.), то бачимо її наявність навіть у Північній Африці.
АФРИКАНСЬКА ГІЛКАR- V88.
Карта R- V88 (Рис. 2.9.) якраз і показує частоти нехай і невисокі (1-6%, з підвищенням до 26% тільки в одній популяції), зате по всій Північній Африці (від Марокко до Єгипту), і майже повна відсутність цієї гаплогрупи в Європі та в Азії. Тобто «африканська» частина R1 b– це майже виключно гілка R- V88 . Свого максимуму вона досягає далеко за межами ареалу, що картографується, - в Центральній Африці.

СТЕПНА ГІЛКАR- M478 .
Карта наступної гілки – R- M478 - Вказує її приуроченість до степових популяцій Євразії (Рис. 2.10). Зона її максимальних частот також знаходиться здебільшого за межами ареалу, що картографується, - на просторах від Уралу до Алтаю. Втім, як можна бачити на карті, окремими мовами ця гаплогрупа доходить і до Кавказу, залишаючись приуроченою до степових областей Південного Уралу і Передкавказзя, і становлячи лише 1-10%, рідко в яких популяціях до 20% генофонду.


ЗАХІДНОЄВРОПЕЙСЬКА ГІЛЛЯR- L265.
Третя з основних гілок гаплогрупи R1 b- Гаплогрупа R-М269,або, як її тут позначаємо на картах, R- L265 — становить основну (за частотою та ареалом) частина R1b. Тому карта її поширення (рис. 2.11) загалом вторить карті L10 лише без африканської частини її ареалу. Ця гаплогрупа поділена на безліч субгаплогруп, багато з яких специфічні для тієї чи іншої території в межах ареалу гілки. Зупинимося на двох основних гілках усередині R- L265 : центральноєвропейській гаплогрупі R- M405 та її піренейсько-британському «браті» R- P312 з усіма їхніми «синівніми» гілками (Рис. 2.12. - 2.17.).

ЦЕНТРАЛЬНОЄВРОПЕЙСЬКА ГІЛКА R-M405 ЦІЛОМ.
Гаплогрупа R-M405(Мал. 2.12) зустрічається практично по всій Західній, Центральній та Північній Європі (за винятком південних областей Піренейського та Апенинського півостровів, а також Балканського півострова – на півдні, Фінляндії та Кольського півострова – на півночі), а також у західній частині Східної Європи . Однак із максимальними частотами (18-36%) гаплогрупа R-M405зустрічається переважно у населенні Центральної Європи (на території Німеччини, Швейцарії, Бельгії, Голландії, Данії та півдні Британії). Датування як виникнення, так і виділення внутрішніх субваріантів гаплогрупи R-M405за даними www.yfull.com становлять близько 5 тис. років тому (в інтервалі 4,4-5,3 тис. років тому), тобто. припадають на бронзовий вік. Схоже, що вже в цей час відбувалися і міграційні процеси, що призводять до поширення гаплогрупи R- M405 у різні частини Європи. Не виключено, що дані про гаплогрупу R- M405 вказують на один із епізодів поширення носіїв індоєвропейських мов на території Європи. На один із напрямів поширення носіїв гаплогрупи R- M405, можливо, вказує географія однієї з її субгілок - гаплогрупи R- M467.

«ГЕРМАНО-БРИТАНСЬКА» ГІЛЛЮR- M467.
Скромне поширення гаплогрупи R- M467 (Рис. 2.13) - як за її частотами (від максимуму 4% у східній Німеччині через значення близько 3% у північній та південній Німеччині, Голландії та на півдні Британії до 0,5% у Франції), так і за географією (не найбільші великі, за європейськими масштабами, території навколо Північного моря) – проте цікавить. Географія гаплогрупи R- M467 наводить на припущення про міграцію її носіїв із території сучасної Німеччини вздовж Північного моря до берегів Британських островів. Датування для гаплогрупи R- M467, представлені на сайті www.yfull.com, вказують на її виникнення близько 4,7 тис. років тому (в інтервалі 4,1-5,4 тис. років тому), а час експансії (поділу на субгілки) через тисячу років – близько 3,4 тис. років тому (в інтервалі від 2,8-4,0 тис. років тому). Порівнюючи ареал гаплогрупи R- M467 на карті із зазначеними датуваннями виникнення та розповсюдження, можна припустити дві моделі появи такої ситуації. Перша – коли міграційні процеси з території центральних областей сучасної Німеччини часів бронзового віку пройшли вздовж берега Північного моря до Британських островів та залишили свій слід у сучасному генофонді. Друга – коли ця гаплогрупа могла бути поширена дещо ширше на південь, захід та схід, але в результаті приходу туди іншого населення і так невисока частота гаплогрупи R- M467 знизилася Півдні до величин, майже вловлюваних при поточних обсягах вибірок.
ПІРЕНЕЙСЬКО-БРИТАНСЬКА ГІЛЛЯR- P312 В ЦІЛОМУ.
Розповсюдження гаплогрупи R- P312 (Мал. 2.14) цікаво тим, що багато в чому відображає загальні особливості розподілу в населенні Європи як своєї предкової гаплогрупи R- L265 , так і всієї гаплогрупи R1 bв цілому. Найвищі частоти гаплогрупи R- P312 на самому заході Європи (Піренейський півострів, територія західної Франції, Британські острови) з поступовим зниженням частоти на схід та локальний сплеск у південному Приураллі – це майже ті ж особливості, що були описані вище при характеристиці ареалу гаплогрупи R1 bв цілому. Однак дотримання загального тренду зовсім не означає повну схожість. Так, велика область червоно-фіолетових відтінків (частоти 50-75%) на території більшої частини Західної Європи, яку спостерігали раніше на картах гаплогруп. R1 b- L10 і R- L265 (Рис. 2.8. та Рис. 2.11.), на карті гаплогрупи R- P312 (Мал. 2.14) перетворюється на локальний сплеск частоти (до 75%) на півночі Британських островів та область жовто-червоних відтінків (частота 30-50%) на території Піренейського півострова та Франції. Південноуральський червоно-фіолетовий сплеск загальної частоти R1 bна карті гаплогрупи R- P312 скорочується до невеликого «вічка» у точці для північних башкир, яку з усіх боків обтікають області низьких частот (трохи більше 3%).
Гаплогрупа R-Р312таким чином охоплює значну частину як різноманітності всієї гаплогрупи. R- L265 , Так її ареалу. Можна припускати, що в такому разі всередині гаплогрупи R- P312 теж існує ряд локальних субгілок з вужчими ареалами. І це припущення виправдовується: на сьогодні відомо п'ять великих гілок, які у свою чергу поділяються на кілька десятків глибших. Однак зупинимося лише на трьох гілках R- P312 , поширення яких у Європі добре вивчено: гаплогрупа R-M167та її субваріант R- M153 , а також гаплогрупа R- M222 (Мал. 2.15-2.17.).

«БАСКО-РОМАНСЬКА» ГАПЛОГРУПАR- M167.
Така назва для гаплогрупи R- M167 умовно: її географія (рис. 2.15) охоплює населення Піренейського півострова і Західної Франції, невелику область Півдні Німеччини і одиничний «вічко» у Голландії, і навіть область у Західному Причорномор'ї (румуни і болгари). Більшість ареалу гаплогрупи, таким чином, населяють народи, що говорять мовами романської (іспанці, португальці, французи та румуни) групи індоєвропейської мовної сім'ї та баскської мовної сім'ї. Втім, південні німці та болгари, у яких теж зустрінуто гаплогрупу R- M167 , говорять мовами інших груп індоєвропейської сім'ї
Область максимальних частот гаплогрупи R- M167 (жовтувато-зелені тони на рис. 2.15, частота гаплогрупи від 6% до 25%) охоплює територію Піренейського півострова та південно-західної Франції. Ця область є єдиною територією поширення дочірньої гаплогрупи. R- M167 - Суваріанту R- M153 (Мал. 2.16).

«БАСЬКА» ГІЛКАR- M153.
Як можна простежити за картою (рис. 2.16), гаплогрупа R- M153 своєї максимальної частоти (майже 16%) досягає у басків, удвічі рідше зустрічається у населенні іспанських Піренеїв, і з дуже низькими частотами (1-3%) охоплює решту свого ареалу. Через таку географію гаплогрупу R- M153 можна умовно назвати «баскською» і припустити, що вона з'явилася та поширювалась переважно у популяціях цього народу. Вік виникнення цієї гаплогрупи за даними www.yfull.com становить близько 2,5 тисячі років (у межах від 3500 до 1500 років тому). Іншими словами, ця гаплогрупа, найімовірніше, з'явилася в популяціях Піренейських гір на зорі нашої ери і встигла поширитися по обидва боки гірського хребта (причому ефективніше – на південь від Піренеїв) внаслідок локальних міграцій.


БРИТАНСЬКА ГІЛКАR- M222.
Дивовижною, тільки не за широтою ареалу, а саме за його компактністю, можна вважати гаплогрупу R- M222 (Рис. 2.17). Поширення цієї гаплогрупи практично виключно на Британських островах наводить на припущення про місцеве її походження. Згідно з оцінками www.yfull.com, експансія гаплогрупи R- M222 датується лише 1,8 тис. років тому (в інтервалі 1,3-2,4 тис. років тому). Іншими словами, гаплогрупа з'явилася в популяціях Британських островів в 2 тис. до н.
Приклад філогеографії гаплогрупи R-L265 показує, як детальна генетична інформація ховається в межах кожної гаплогрупи Y-хромосоми. Залишається сподіватися, що в міру накопичення даних по геномах будуть виявлені інформативні маркери і в межах інших гаплогруп.
ІНДІЙСЬКА ГАПЛОГРУПАR2 (R- L266).
Гаплогрупа R2 (R- L266 ) походить з того ж кореня, що і гаплогрупа R1 , що дала початок гаплогрупам R1 aі R1 b. Карта поширення гаплогрупи R2 (R- L266 ) (рис. 2.18) показує, що для населення Європи в цілому R2 не характерна: вона зустрічається з вкрай низькими частотами (менше 3%) лише на півночі Апенінського півострова та в Сардинії, біля популяції турків поблизу протоки Дарданелли, у центральній Анатолії та Закавказзі. З трохи вищою частотою 5% гаплогрупа R2 (R- L266 ) помічена в південно-східній Туреччині, із частотою 6% - у калмиків. Така мізерна географія гаплогрупи в межах ареалу, що картографується, мабуть, не дивна: адже основний ареал R2 (R- L266 ) охоплює далекі території Індійського півострова та Центральної Азії, а з низькими частотами гаплогрупа доходить до Китаю на сході та до Південно-Західної Азії – на заході свого ареалу.

Продовжуючи тему про високу інформативність виділення субтипів та аналізу їх географічного розподілу, нагадаю класичну роботу, що стала гаплогрупою. I-М170була філогенетично поділена на три субтипи. Виявилося, що ці субтипи мають зовсім різну географічну приуроченість : гаплогрупа I1-М253(на картах - I-L118) присвячена північній Європі, I2a-Р37– до Балканського регіону, а I2b-М223(на картах - I-L35) локалізована переважно у Північно-Західній Європі. Саме після цієї роботи та ряду їй подібних стало ясно, що виявлення нових SNP маркерів, що підрозділяють гаплогрупу на субгілки, є магістральним шляхом підвищення інформативності аналізу Y-хромосоми. Але довгий час рух цим шляхом був повільним, і лише після впровадження повного секвенування Y-хромосоми нові маркери стали накопичуватися лавиноподібно. Так, протягом тривалого часу не вдавалося знайти нові SNP маркери в межах гаплогрупи I, або нові SNP маркери не виділяли нових гілок з чіткою географією. Наприклад, у роботі було виявлено нові маркери та реорганізовано топологію гілок гаплогрупи. I-М170, але збереглися, хай під новими назвами, самі три основні гілки, а нові виявлені варіанти були рідкісними і характеризувалися чіткими географічними трендами.
«СКАНДИНАВСЬКА» ГАПЛОГРУПА I-L118.
Карта поширення «скандинавської» гаплогрупи I-L118(Мал. 2.19) показує її максимальні частоти у географічної Скандинавії, а й у територіях, тісно пов'язаних із нею історично: Данії, Шотландії, західних районах Фінляндії. Тому цікава область помірних частот у російських популяцій на північ від Волги, що виділяється на тлі низьких частот у більшості слов'янських популяцій. Не можна виключати, що це відбиває історичні контакти з норманськими населеннями. Хоча ці контакти були інтенсивніші для північно-західних російських популяцій (по дорозі «з варяг у греки»), але території північного заходу були порівняно густо населені, тому вплив «варягів» міг бути більш значним для менших за обсягом популяцій Заволжя.

«БАЛКАНСЬКА» ГАПЛОГРУПА I-Р37
Карта поширення «балканської» гаплогрупи I-Р37(Мал. 2.20) показує її високі частоти й у Італії. Загалом ця гаплогрупа поширена практично по всій Європі, крім Скандинавії. У Східній Європі плавний градієнт зменшення її частоти з південного заходу (з України) на північ і схід дуже нагадує карту першої головної компоненти з класичних генетичних маркерів [Ричків та ін., 2002]. Також варто відзначити досить виражений збіг ареалу цієї гаплогрупи з географічними кордонами Європи – за високої частоти в Причорномор'ї гаплогрупа дуже рідка на Північному Кавказі і майже відсутня у Закавказзі, а за максимальних частот на Балканах дуже рідко в сусідній Малій Азії.

«ВАРЯГО-грецька» ГАПЛОГРУПА I-L35
Уздовж берегів Північного і Балтійського морів, і трохи вглиб від них так можна було б описати географію гаплогрупи I-L35, якби ще не два доповнення на північному сході та південному сході. Хоча максимум частоти гаплогрупи I-L35 ( 17%) відзначений у Швеції (рис. 2.21), зона більш рідкісної гаплогрупи (3-7%) охоплює північну частину Центральної Європи, на південному сході окремою областю дуже низьких частот (1-3%) охоплює Балканський півострів і західну частину Малої Азії, але в північному сході проявляється в російських популяцій Тверської і Костромської областей. Можна помітити, що ареал гаплогрупи I-L35 — досить помірний і однорідний за значеннями частот гаплогрупи (на більшості ареалу частота варіює від 2-3% до 7%, лише пік у Швеції до 17%) - пов'язує кілька крайніх областей Європи: Скандинавію, північні райони Східної Європи та схід Південної Європи. Цілком можливо, що географія гаплогрупи I-L35 дійсно відображає один із генетичних слідів торгових шляхів між північчю та півднем Європи. Не менш імовірно, що кожна із цих зон пов'язана зі своїм субваріантом цієї гаплогрупи.

Гаплогрупа Nу межах Європейського регіону представлена двома великими гілками - гаплогрупами N1cі N1b(далі і на картах N-M46і N-P43відповідно). Хоча загальні ареали цих двох гаплогруп у Європі перетинаються, зони їх максимальних частот розмежовуються на півночі річкою Печорою, а на південь — Уральським хребтом: із заходу цієї умовної «кордону» залишаються зони підвищеної гаплогрупи N-M46, а зі сходу - зона максимальних частот N-P43. Розглянемо географію цих двох «братських» гілок гаплогрупи Nтрохи докладніше.
ПІВНІЧНО-ЄВРОПЕЙСЬКА ЗАХІДНА ГАПЛОГРУПА N-M46
Карта поширення гаплогрупи N-М46(Мал. 2.22) показує її максимальні частоти у фінів. Її частоти високі та в інших популяцій північного сходу Європи. Хоча в цілому ця гаплогрупа виявляється приуроченою до фінно-угорських популяцій, але вона така ж часто й у північних росіян (що, мабуть, пояснюється асиміляцією дослов'янського населення, як буде описано в наступних розділах). Найбільш цікавою є область високої частоти цієї гаплогрупи у балтів (латишів і литовців). У разі генетичні кордону добре збігаються з лінгвістичними: кордон ареалів балтів і західних слов'ян збігається з різко вираженим «обривом» частоти гаплогруппы. Схожа картина спостерігається на кордоні з білорусами.

«ПІВНІЧНО-УРАЛЬСЬКА» ГАПЛОГРУПА N-P43
Карта поширення гаплогрупи N-P43(Мал. 2.23) виявляє також північний, але куди вужчий ареал. Максимальні частоти цієї гаплогрупи спостерігаються на крайньому північному сході Європи та плавно переходять у світовий максимум цієї гаплогрупи у Західному Сибіру. З помірними частотами ця гаплогрупа поширена по всьому Уралу та Пріураллю. Помітний плавний градієнт зменшення частоти на захід і південь, так що у популяцій Центральної Росії та нижньої Волги ця гаплогрупа ще зустрічається (хоча і з мінімальними частотами), а в інших популяцій Європи практично повністю відсутня.

Більшість вищеописаних гаплогруп переважно характеризували умовно північну половину Європейського континенту. Розглядаючи географію наступної гаплогрупи E– ми перемістимо увагу на південь до берегів Середземного моря. У Європі зустрічаються чотири основні гілки гаплогрупи E: E- L142 і E- V22 (висхідні до загального кореня - гаплогрупи E-M78), E- M81 і E- M123 .
СХІДНО-СРЕДИЗЕМНОМОРСЬКА ГАПЛОГРУПАE- L142
Карта гаплогрупи E- L142 (Мал. 2.24) частково нагадує карту гаплогрупи I-Р37(Рис. 2.20) - максимальні частоти обох цих гаплогруп приурочені до Балканського півострова. Також дуже схожі їхні патерни: зменшення частоти на північ, захід і схід. Основні відмінності між трендами двох цих гаплогруп складаються, по-перше, у меншій частоті E- L142 (На більшості територій вона «поступається» I-Р37на один інтервал (шкали карти) і, по-друге, у поширенні E- L142 не тільки в Європі, а й по всьому Середземномор'ю, включаючи Близький Схід та північні околиці Африки. У межах Балкан I-Р37особливо часто у боснійців та хорватів, а E- L142 переважає південніше – у сербів, албанців та греків.

«ЄГІПЕТСЬКА» ГАПЛОГРУПАE- V22
Два невеликі «очі» на півночі та півдні Піренейського півострова та Апенінський півострів – території з частотою гаплогрупи E- V22 до 5%, невелика область на північному заході Малої Азії (на околицях Стамбула) з частотою до 6%: на цьому можна було б закінчити опис географії гаплогрупи E-V22 у географічних межах Європи (рис. 2.25). Проте географія гаплогрупи E-V22 в Європі в основному слідує Середземноморському узбережжю, тому варто простежити її поширення і на південь. Мапа на рис. 2.25 показує плавне підвищення частоти гаплогрупи E- V22 у двох протилежних регіонах: на південному заході та південному сході Середземномор'я. У південно-західному Середземномор'ї біля берегів Гібралтарської протоки та вглиб Марокко частота гаплогрупи E-V22 зростає до 7%. У південно-східному Середземномор'ї підвищення частоти гаплогрупи E- V22 відзначається від Малої Азії на південь - через Левант аж до дельти Нілу, де вона досягає 14%, і ще південніше вздовж Нілу, де у єгипетських арабів оази Бахарія вона досягає світового максимуму 22%. З цих двох неєвропейських регіонів поширення гаплогрупи E- V22, ймовірно, саме останній (з вищими частотами та широким поширенням) послужив основним джерелом поширення гаплогрупи E- V22 в Європу.

САХАРСЬКА ГАПЛОГРУПАE- M81
Якщо дві попередні гілки гаплогрупи E – E- L142 і E-V22 – демонстрували в основному поширення вздовж північних та східних берегів Середземного моря, іноді з поглибленням на північ та схід континенту, то географія третьої гілки – гаплогрупи E- M81- більше слід південному Середземномор'ю. Карта поширення гаплогрупи E- M81 на рис. 2.26 хоч і показує помітну присутність гаплогрупи на Близькому Сході (частота до 20%), проте основна зона високих частот знаходиться у північно-західній Африці. З цього максимуму зазначено плавне зниження частоти в північному напрямку через Гібралтарську протоку з помірним поширенням (2-10%) гаплогрупи E- M81 на території Піренейського півострова та південно-західної Франції, вузькі області низьких частот (не більше 3%) на Аппенінському та Балканському півостровах, а також на північному сході Малої Азії.


ПЕРЕДНЕАЗІАТСЬКА ГАПЛОГРУПАE- M123
Гаплогрупа E- M123 досягає максимуму в Передній Азії (20-25%), ближче до Європи (у Малій Азії) знижується до 15%. У самій Європі вона з низькими частотами поширена вздовж північних берегів Середземного моря, і окремими вкрапленнями проявляється у Центральній Європі (рис. 2.27).
Карти розповсюдження гаплогрупи двох гілок гаплогрупи G- P15 – гаплогруп G- P303 таG- P16 – загалом мають схоже поширення і навіть у загальноєвропейському масштабі карток видно, що обидві приурочені насамперед до Кавказу. Однак своїх максимальних частот вони досягають у різних частинах Кавказу регіону: гаплогрупа G- P303 переважає на крайньому північному заході у причорноморських шапсугів, а гаплогрупа G- P16 - на Центральному Кавказі в осетин-іронців (рис. 2.28 та рис. 2.29). Це повністю підтверджується при окремому вивченні генофонду Кавказу.

ЗАХІДНОКАВЯЗЬКА ГАПЛОГРУПАG- P303
Карта показує, що гаплогрупа G- P303 (Мал. 2.28) також поширена на Близькому Сході, а в Європі зустрічається в основному в її південно-східних областях. Зони високої частоти гаплогрупи G- P303 тягнуться вздовж східного узбережжя Чорного моря і присвячені популяціям народів Західного Кавказу, переважно, абхазо-адизької лінгвістичної групи. З меншими частотами (до 10%) гаплогрупа G- P303 поширена у Північному Причорномор'ї (на території Кримського півострова та у Приазов'ї), область низьких частот (3-5%) тягнеться далі на північ у зону українського лісостепу. Локальне підвищення частоти гаплогрупи G- P303 до 13% також спостерігається у Поволжі у популяції мордви мокша та оточується областю низьких частот.

ЦЕНТРАЛЬНОКАВ'ЯЗЬКА ГАПЛОГРУПАG- P16
Географія гаплогрупи G- P16 (Рис. 2.29) Вужча в порівнянні з її «братської» гілкою G- P303 . Вона охоплює виключно Кавказький регіон та Малу Азію. У межах цього ареалу частота гаплогрупи G- P16 різко падає від максимальних значень у осетин (73% у осетин-іронців і 56% у осетин-дигорців) до значень в діапазоні 10-20% у народів Північно-Західного Кавказу (абхазів, черкесів, балкарців та карачаївців) і потім знижується до мінімальної позначки у турків, вірмен та азербайджанців.
Гаплогрупа Jпредставлена двома великими гілками: J- L255 (J1) і J-L228 (J2),з яких у Європі більшого поширення набула друга. Хоча обидві ці гілки за своїм походженням близькосхідні і відображають стародавні взаємозв'язки населення Європи та мешканців набагато південніших регіонів Євразії, має сенс розглянути окремо географію і кожної з цих ліній загалом, і кожної із внутрішніх гілок більш поширеної в Європі гаплогрупи. J-L228 (J2).

ГАПЛОГРУПАJ- L255 (J1): від Передньої Азії до вершин Східного Кавказу
Розповсюдження гаплогрупи J- L255 (Мал. 2.30) в ареалі, що картографується, охоплює переважно його південно-східні області: Кавказ, Малу Азію, Північне Причорномор'я, Балканський півострів; невеликими зонами – південь Апенінського півострова та Франції, південний захід Піренейського півострова. У межах регіону максимальних частот – Кавказу – гаплогрупа J- L255 виділяє народи Східного Кавказу, де її частку припадає левова частка всього генофонду. Максимума гаплогрупа J- L255 досягає в Дагестані (у кубачинців, даргінців, табасаранців та аварців її частоти коливаються від 63 до 98%), і знижується до 44% у лезгін та 21% у чеченців. На території Малої Азії частка гаплогрупи J- L255 становить від 5% до 15%.
ГАПЛОГРУПАJ-L228 (J2) В ЦІЛОМУ
Карта поширення гаплогрупи J-L228 (J2)малює іншу картину (рис. 2.31). На відміну від більшості розглянутих вище гаплогруп, J-L228 (J2)поширена з високими частотами над Європі, але в Близькому Сході й у Північній Африці. Звичайно, її ареал заходить і в сусідні з цими регіонами південні області Європи: Іспанію (особливо її південні райони), Італію (також особливо південну), південь Балканського півострова. Але максимальних частот (червоний колір на карті) ця гаплогрупа досягає деяких популяціях Північного Кавказу. Як було показано в нашому дослідженні, а також у роботі кавказький максимум присвячений популяціям нахської групи (чеченцям та інгушам).


ГАПЛОГРУПАJ- L152 І ЇЇ СУБВАРІАНТJ- M67
Географія гаплогрупи J- L152 на території Європи досить велика: зона низьких частот тягнеться з північного заходу Балканського півострова через Центральну Європу та до Піренейського півострова. Зона підвищених частот (від 15%) охоплює Малу Азію та Кавказ. Практично повторює цю закономірність, але дещо у вужчому географічному охопленні й загалом із нижчими частотами, поширення її «синівньої» гілки – гаплогрупи J-M67(Рис. 2.32). Однак при цьому гаплогрупа J-M67за своєю частотою більше присвячена Кавказу, складаючи біля Малої Азії трохи більше 13% генофонду.
ГАПЛОГРУПАJ- L282
Розповсюдження гаплогрупи J- L282 у Європі (рис. 2.33) помітно відрізняється від попередньої гілки. На тлі великої зони низьких частот виділяються три області, де частка гаплогрупи J- L282 у генофонді підвищено до 10-15%: північ Фенноскандії (щоправда, цей максимум спирається лише дані по одній популяції), Поволжя (популяція мордви мокша) і північ Балканського півострова (кілька популяцій албанців).

РІДКІ ГАПЛОГРУПИL, Q, T
ГАПЛОГРУПА L-M11
Гаплогрупа L-M11(рис. 2.34) у Європі приурочена до південно-східних регіонів (за винятком поодинокого випадку в Бельгії): Кримський півострів, європейська частина Туреччини та Північний Кавказ. При цьому частота гаплогрупи L-M11плавно збільшується у південно-східному напрямі, досягаючи 14% біля Східного Кавказу в чеченців і 13% північному сході Малої Азії в турків. Далі її частота підвищується на Близькому та Середньому Сході, досягаючи індійського максимуму вже далеко за межами ареалу, що картографується.

ГАПЛОГРУПА Q-M242
Гаплогрупа Q-M242у Європі практично не зустрічається, за винятком кількох точок з низькими частотами (до 3%) у Центральній та Східній Європі, на Кримському півострові та у східній половині Малої Азії (рис. 2.35). Плавним коридором від північно-західного узбережжя Каспійського моря на Схід з поступовим зростанням частоти ареал гаплогрупи Q-M242переходить у Центральну Азію та Сибір, де вона досягає свого максимуму.


ГАПЛОГРУПА T
Частота гаплогрупи T-L206у народонаселенні Європи не перевищує 5% (рис. 2.36). Її ареал у Європі клаптевий і зводиться на заході до трьох вузьких областей на Піренейському півострові та в Сардинії, а на сході – до зони в Північному Причорномор'ї з переходом на північний схід в український лісостеп, поодинокими випадками в Поволжі та вказаних у кубанських ногайців За межами Європи частота гаплогрупи Т настільки ж невелика, зате її ареал стає вже не клаптевим, а майже безперервним, охоплюючи всю Малу Азію, Близький Схід та північний схід Африки.
Таким чином, проведене дослідження мінливості Y-хромосоми в популяціях Європи, засноване на об'єднанні великих власних даних з літературними, підтвердило та уточнило висновок про те, що основною рисою структурованості генофонду є чітке виділення географічних зон, у кожній із яких переважає своя гаплогрупа. Це переконливо демонструють карти розподілу гаплогруп (рис. 2.2-2.36).
Однак при розміщенні даних по кожній гаплогрупі на окремій карті важко відстежити перекриття - або, навпаки, не перекриття - ареалів гаплогруп. Тому ми поєднали зони поширення всіх гаплогруп на одній карті (рис. 2.37). На цьому малюнку контур кожної з дев'яти основних європейських гаплогруп суворо відповідає території, де частота цієї гаплогрупи вище 35%, тобто. понад третину генофонду. Можна бачити, що кожна гаплогрупа справді займає свій власний ареал. Області, що залишаються білими, невеликі – це території, на яких різноманітність гаплогруп високо і жодна гаплогрупа не досягає рівня 35%.
Отримана карта яскраво демонструє "принцип пазлів" - високу географічну специфічність гаплогруп Y-хромосоми. Завдяки цьому на кожній території, хоч і є багато гаплогруп, але домінують лише одна-дві. Таким чином генофонд Європи виявляється складений, як із пазлів, із ареалів переважного поширення різних гаплогруп.

У кожній географічній частині Європи домінує одна гаплогрупа, яка рідко зустрічається в інших частинах. Карта показує, до яких частин Європи приурочено кожну з основних гаплогруп. Карта ґрунтується на точних значеннях частот. Кольором показані зони з частотою гаплогрупи вище за порогове значення 0,35 (тобто на виділених кольором територіях більше третини генофонду відноситься до даної гаплогрупи).
Зазначимо в дужках, що об'єктивна приуроченість даної гаплогрупи до слов'янських популяцій на цій дуже обмеженій території, на жаль, стала одним із приводів настільки поширеного на аматорських форумах в інтернеті навішування «слов'янського» ярлика на всю гаплогрупу R1a. Але така прив'язка гаплогрупи — на всьому протязі її євразійського ареалу в просторі і несумірного зі слов'янством тривалого існування в часі — лише до однієї слов'янської лінгвістичної групи, на жаль, уже не тільки межує з лженаукою, а й переходить цю грань.
Якщо серед сучасних європейських народів Європи можна виділити кілька основних гаплогруп (понад 5%), це звичайно ж R1b, R1a, I1, N1, I2, G2 на півночі та J, E1b1, T на півдні Європи, то щодо населення стародавньої Європи ситуація зовсім інша.
Субклад I2-M26 це одна з основних Y-хромосомних ліній на Сардинії (досягає 40%) та у басків. З низькою частотою зустрічається також у всій південно-західній Європі: Іспанії, Франції, Португалії, Італії.
Субклад I2-M423 характерний для населення Балкан та Карпат, найчастіше спостерігається у динарійських слов'ян (серби, хорвати та боснійці), а також у Молдові та на південно-західній Україні. Також з помітною частотою виявляється в Албанії, північній Греції, Болгарії, Словаччині, на східній Україні, Білорусії та південно-західній Росії.
Субклад I2-M223 характерний для північно-західної Європи та досягає максимуму в Німеччині (Нижня Саксонія). Також зустрічається в Румунії, Молдові та з низькою частотою у Східній Європі.


Дані гаплогрупи з різною частотою від 0-95% зустрічаються серед німців, іспанців, італійців, греків, поляків, угорців, французів, албанців, англійців, ірландців, шотландців, португальців, швейцарців, чехів, словаків, сербів, хорватів, румунів українців, датчан, шведів, норвежців, фінів, росіян, естонців, латишів, литовців та ін.
У 80-ті роки ХХ століття у зв'язку з вивченням структури генетичного коду відбулася "безшумна революція" в антропології. Виникла нова галузь науки, що називається палеогенетикою чи молекулярною палеонтологією. Виявилося, що у самій людині, точніше - у її генотипі, що є сукупністю всіх генів організму, можна знайти сліди еволюційної історії виду. Гени вперше постали в ролі надійних історичних документів, з тією лише різницею, що запис у них зроблено не чорнилом, а хімічними компонентами молекули ДНК. Генетики навчилися витягувати інформацію в буквальному значенні з «праху земного» - закам'янілих залишків, які належали дуже давнім істотам. Отримані палеогенетикою дані радикальним чином трансформували попередні уявлення про ранні стадії «людської» еволюції.
Мітохондріальна Єва - ім'я, дане молекулярними біологами жінці, яка була останнім спільним предком всіх людей, що нині живуть по материнській лінії. Оскільки мітохондріальна ДНК успадковується тільки по материнській лінії, у всіх людей, що нині живуть, така ДНК була отримана від «Єви». Аналогічно ДНК чоловічої Y-хромосоми у всіх людей чоловічої статі має походити від "молекулярно-біологічного Адама".
На відміну від ядерної ДНК, яка містить переважну більшість генів і в процесі статевого розмноження піддається рекомбінації, так що нащадки отримують половину генів від батька, а другу половину від матері, мітохондрії та їх ДНК дитина отримує тільки з материнської яйцеклітини. Оскільки мітохондріальна ДНК не піддається рекомбінації, зміни в ній можуть відбуватися виключно за допомогою рідкісних випадкових мутацій приблизно один раз на 3000 років. Шляхом порівняння послідовності мітохондріальної ДНК і виникли в ній з часом мутацій можна не тільки визначити ступінь спорідненості людей, що нині живуть, а й приблизно обчислити час, необхідний для накопичення мутацій в тій чи іншій популяції людей.
Провівши порівняльний аналіз мтДНК, 1980 року А. Вілсонпобудував генеалогічне дерево, яке чітко свідчило про наявність найбільшої диференціації генів мітохондріальних в Африці. Більше того, все шестимільярдне сучасне людство веде своє походження від однієї жінки, яка колись мешкала у східній Африці, оскільки всі досліджені зразки мтДНК можна звести до єдиної вихідної нуклеотидної послідовності. А. Вілсон, знайшовши місце, що є «колискою» людства, пішов далі. Знаючи швидкість мутування, він зміг визначити і приблизний час, коли Єва з'явилася на Землі. «Мітохондріальний годинник» показав, що вона жила приблизно 200-150 тисяч років тому (дивно, але «Єва» виявилася навіть давнішою за неандертальця, якого наполегливо нав'язували їй в «еволюційні батьки»).
Дані щодо аналізу мтДНК були незалежно отримані багатьма іншими дослідниками. «Аналіз мтДНК, – пише Сатосі Хораї,- вказує на те, що сучасна людина виникла близько 200 тисяч років тому в Африці, звідки переселилася до Євразії, де досить швидко витіснила Homo erectus і, ймовірно, повністю (якщо не буде знайдена снігова людина) неандертальця. При цьому змішування мітохондріальних генотипів практично не сталося.
1987 року Ребекка Канн (Rebecca Cann)з колегами припустили, що мітохондріальна Єва могла жити між 140 та 280 тис. років тому. Згідно з пізнішими розрахунками 2004 р., мітохондріальна Єва жила близько 140 тис. років тому в Східній Африці. Сучасні МП- та МЕ-оцінки 2009 р. зазвичай дають діапазон віку Єви 140000-230000 років, з максимумом ймовірності на значеннях порядку 180000-200000 років.
Особливий інтерес представляє Л. Каваллі-Сфорцаспроба порівняти дані молекулярної генетики та лінгвістики. Він показав, що поширення генів напрочуд добре корелює з поширенням мов. Таким чином, родовід дерево, побудоване на підставі генетичних досліджень, відповідає лінгвістичному родоводу. Так геногеографія поєдналася з етнічною географією.
Ще за життя А. Вілсона була зроблена спроба аналізу У-хромосоми чоловіків з тим, щоб простежити «лінію батьків» у родоводі людства. Попередні дані, про які він повідомляє, отримані французьким ученим Ж. Люкоттом, також підтвердили африканське походження Адама.
Більш детальні дослідження провели професор Стенфордського університету. П. Ундерхаллом,зібравши матеріал для аналізу майже у всіх регіонах світу. Як відомо, У-хромосома присутня лише в генотипі чоловіків, і, отже, передається у поколіннях строго від батька до сина. Результат вивчення кількох тисяч проб, взятих від представників різних народностей, показав, що батьківщиною «Адама» була та сама східна Африка. За оцінкою дослідників, час появи представника Homo sapiens чоловічого роду становить близько 150-160 тисяч років. Деякий розкид у віці «Єви» та «Адама» укладається у межі помилки методу.
Аналогічні дані було отримано й іншою незалежною групою під керівництвом Майкла Хаммера(Університет Арізони, США). Уточнений вік гіпотетичного «Адама» – 160-180 тисяч років.
Отже, саме на Африканському континенті близько 150-200 тисяч років тому з'явилися наші прабатьки. Приблизно 100 тисяч років тому їхні нащадки мігрували по всій ойкумені, заміщаючи всіх інших гомінід, що жили там, але при цьому, що важливо, не схрещуючи з останніми. Близько 40-60 тисяч років тому вони дісталися Європи.
Але на цьому сюрпризи, подані палеогенетиками антропологам, не закінчилися. Професору Сванте Паабовдалося витягти мтДНК із фрагмента хребця неандертальця, вперше виявленого в 1856 р., який жив близько 50 тисяч років тому. Ця робота - справді висока вершина молекулярно-генетичного мистецтва, результат її важко переоцінити. Як показали порівняльні дослідження мітохондріальної ДНК сучасної людини та неандертальця, останній зовсім не є ні нашим предком, ні навіть близьким родичем. Шляхом порівняльного аналізу «наших» і «неандертальських» генів було встановлено, що різницю між ними настільки великі, що еволюційні гілки цих двох видів могли (чи мали) розійтися 600 тисяч років тому, тобто у той час, коли самих видів ще просто не існувало.
Висновки С. Паабонастільки докорінно змінюють уявлення про антропогенез, що постало питання про перевірку цих результатів незалежною групою дослідників. Із фрагментом неандертальської кістки цього разу працював Марк Стоункінґ,вчений із групи А. Вілсона,також високий авторитет у галузі палеогенетики. Провівши дослідження мтДНК з іншого зразка (останки неандертальської дитини, яка жила 30 тисяч років тому), вона отримала такі ж дані, що й С. Паабо,повністю підтвердивши його висновки. У зв'язку з цим в одному з інтерв'ю С. Паабо зауважив: «Ми дотримувалися суворих критеріїв судової медицини, ніби готувалися подати речовий доказ суду».
Через кілька років група німецьких учених також провела незалежне дослідження неандертальської мтДНК, яке показало: «Підтверджується гіпотеза, згідно з якою неандертальці представляють еволюційну тупикову гілку і не є предками сучасної людини» .
Палеонтолог Крістофер Стрінгертак бачить подальшу перспективу: «Можливо ми стоїмо на порозі створення єдиної теорії, яка об'єднає палеоантропологічні, археологічні, генетичні та лінгвістичні докази на користь Африканської моногенетичної моделі» .
Справді, синтез цих наук, мабуть, здатний наблизити нас розуміння таємниці нашого походження.
Дерево гаплогруп мтДНК людини
| Мітохондріальна Єва | |||||||||||||||||||||||||
| | | |||||||||||||||||||||||||
| | | |||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||
| | | | | | | | | ||||||||||||||||||||||
| pre-JT | N1a | ||||||||||||||||||||||||
| | | | | | | |||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||
Дерево гаплогруп Y-ДНК людини(Гаплогрупи Y-ДНК за народами)
| A1b | A1a-T | |||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A2-T | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2 | A3 | |||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ||||||||||||||||||||||||||||||||||||||||||||||||||
| K | ||||||||||||||||||||||||||||||||||||||||||||||||||
| K(xLT) | ||||||||||||||||||||||||||||||||||||||||||||||||||
Розглянемо K- Y-хромосомну гаплогрупуз наступними субкладами (L, T, M, NO, P та S). К - Y-хромосомна гаплогрупа з'явилася 40-50 тис. років тому. ймовірно в Передній Азії.
За останніми даними нащадки:

Розповсюдження гаплогрупи L.
Середньостатистичний індус
. Для гаплогрупи L спостерігається висока частота та різноманітність підкласів на південному заході Пакистану в Белуджистані вздовж узбережжя (28 %). Національний різновид виник 24-30 тис. років тому.
Папуас. Національний різновид виник 32-47 тис. років тому.
Папуаси Ekari. Національний різновид виник 28-41 тис. років тому.
Розповсюдження гаплогрупи T.
Фульбе - народ, який проживає на широкій території в Західній Африці: від Мавританії, Гамбії, Сенегалу та Гвінеї на заході до Камеруну і навіть Судану на сході. Для гаплогрупи T спостерігається висока частота та різноманітність підкласів (18 %). Національний різновид виник 19-34 тис. років тому.
- N гаплогрупа.Ця гаплогрупа зустрічається у Центральній, Північній Європі та повсюдно у європейській та азіатській частинах Росії. Найбільш генетично «чисті» представники - якути (74%), ненці (74%), удмурти (68%), фіни (61%), народи уральських мов, ескімоси. Переважна більшість сучасних представників цієї групи належать до гілки N1, найбільш ймовірне місце появи якої - регіон Алтаю, Прибайкалля, південного Сибіру, Монголії або північного Китаю, а час - між 20 000 і 15 000 років тому. Вважається, що вона була принесена через Євразію великою міграцією лісових сибірських народів на захід, остання фаза якої – розселення по Уралу і далі з нього по Поволжі та північно-східній Європі (Російська Північ, Фінляндія, Прибалтика) – асоціюється з поширенням у цьому регіоні уральських мов.
Ненці – самодійський народ, що населяє євразійське узбережжя Північного Льодовитого океану від Кольського півострова до Таймиру. Для гаплогрупи T спостерігається найвищий показник (74%) . Національність виникла 15-20 тис. Років тому.
- Гаплогрупа O- Y-хромосомна гаплогрупа людини, що є нащадком гаплогрупи Гаплогрупа NO, вперше з'явилася відповідно до різних теорій, або в Південно-Східній Азії або Східній Азії 28-41 тис. років тому; характерна для представників монголоїдної раси. Родична «фінно-угорській» гаплогрупі N. Ця гаплогрупа з'являється в 80-90% більшості населення в регіоні Східної та Південно-Східної Азії у китайців, японців, філіппінців, малайців, австронезійців, а також у суміжних народів, на яких вони вплинули субстрату. Цілком відсутня ця гаплогрупа в Європі, Західному Сибіру, Близькому Сході, Африці та Америці. Представляє близько 21% усієї глобальної людської популяції.
Розповсюдження гаплогрупи O.
Середньостатистичний китаєць - включає всі 56 груп, що проживають у Китаї та офіційно визнані урядом, наприклад, такі як монголи, маньчжури, тибетці, та інші встановлені етнічні групи, які проживають у Китаї хоча б з династії Цін (1644—1911). Для гаплогрупи T спостерігається висока частота та різноманітність підкласів,найвищий показник (60-80%). Національність виникла 28–41 тис. років тому.
Реконструкція фізичного вигляду неоантропів-європеоїдів Російської рівнини епохи верхнього палеоліту:
зліва сунгірійка (поселення Сунгір, м. Володимир, бл. 30 тис. л.н.), праворуч - костенківець (поселення Костенки, Воронезька обл., бл. 45 тис. л.н.). Реконструкції М.М. Герасимова.
Дана гаплогрупа розбивається на субклади Q та R:
- Q гаплогрупа,час появи 15-20 тис. років до зв. е.., поширена в деяких сибірських народів, а також у корінних американських народів, і, певною мірою - по всій Азії. Передбачається, що носіями даної гаплогрупи були гуни, що мають сибірське походження. В Євразії зустрічається в межах трикутника з вершинами в Норвегії, Ірані та Монголії. Але здебільшого серед усіх цих народів зустрічається рідко. У Європі дана гаплогрупа поширена серед угорців (2%) та словаків (5%). Однак значна у нечисленних сибірських народів кети (95%) та селькупи (70%). Типова й у корінних американців.
Сім'я кетів чи єнісейців.
Антропологія схожа на корінних індіанців і народів Океанії, наприклад, полінезійців (Нова Зеландія). Це можливий образ людини цієї гаплогрупи протягом 15-20 тис. років.
- R гаплогрупа,гаплогрупа R зародилася між 30 000 та 35 000 років тому. На відміну від Q зазнає змін і має субклади, що вказує на її давнину. Цілком імовірно, що саме до групи IJналежали кроманьйонці (або їх більша частина), перші представники
Поширення гаплогрупи R по субкладах R1a (бузковий) та R1b (червоний).
- Гаплогрупа R2— зустрічається вкрай рідко, переважно на території Ірану, Індії, Пакистану, Північного Кавказу.
- Гаплогрупа R1— найпоширеніша підгрупа Гаплогрупи R. Її два основні субклади R1a і R1b (інші варіанти зустрічаються виключно рідко) є найпоширенішими у всій Європі та західній Євразії. Це з переселеннями після останнього льодовикового максимуму. Передбачається, що гаплогрупа R1 могла зародитись 25,000-30,000 років тому.
- Гаплогрупа R1a,ймовірно, зародилася Півдні Російської рівнини приблизно 10-15 тис. років тому вони. Вважається, що на їх основі сформувався, зокрема, слов'янський етнос. Ареал розповсюдження від Ісландії до Індії, сучасний центр гаплогрупи знаходиться на території Польщі. Ця гаплогрупа стала маркером поширення курганної культури, яку, своєю чергою, більшість авторитетних дослідників нині вважають ядром протоіндоєвропейської культури (Курганна гіпотеза). Генетики показали, що курганні поховання скіфськихскелетів містять гаплогрупу R1a. Експансія сприяла міграції гаплогрупи R1a до Ірану та Індії, де близько 30 % чоловіків у вищих кастах є її носіями. Найбільшого поширення має Східна Європа: серед лужичан (63 %), поляків (бл. 56 %), росіян (53 %), білорусів (49 %), українців (бл. 52 %), татар (34 %), башкир ( 26%) (у башкир Саратовської та Самарської обл. до 48%); та в Центральній Азії: у худжандських таджиків (64 %), киргизів (63 %), ішкашимців (68 %). Помірне поширення в Скандинавських країнах (23% в Ісландії, 18-22% у Швеції та Норвегії), в Ірані (25%?). У брахманів індійських штатів Західна Бенгалія та Уттар-Прадеш дана гаплогрупа зустрічається з частотою 72% та 67% відповідно.
Середньостатистичний поляк. Для гаплогрупи R1a спостерігається найвищий показник (56%).
- Гаплогрупа R1b, Різні автори відносять появу даної гаплогрупи до 16 - 5 тис. років до н. е.; поширена найбільшою мірою на заході Європейського континенту, особливо високий відсоток її носіїв серед народів західної Англії, басків, башкир на Південному Уралі та іспанців. У Східній Європі та на сході Гаплогрупа R1b зустрічається у вірмен, у Дагестані. Носієм був фараон Тутанхамон. Сучасна концентрація R1b максимальна на територіях у південній Англії близько 70 %, у північній та західній Англії, Уельсі, Шотландії, Ірландії – до 90 % і більше, в Іспанії – 70 %, у Франції – 60 %, басків – 88,1 % та іспанців - 70%. у італійців - 40%, німців - 39%, норвежців - 25,9%
Середньостатистичний англієць. Для гаплогрупи R1b спостерігається найвищий показник (близько 70%).Національність виникла 10-15 тис. Років тому.
Для повної картини народження гілок сучасних національностей до описаної вище гаплогрупі Ктак само слід виділити інші, які визначають чи посилюють належність до національності.
Поряд із гаплогрупою Кна кілька тисяч років після виникає гаплогрупаIJ (і її нащадки I і J ) . Час появи 38,5 (30,5-46,2) тис. років тому. Цілком ймовірно, що саме з'єднанню групі IJ і Р (з нащадками R) належали кроманьйонці (або їх велика частина), перші представники людей сучасного типу європеїду, що прийшли до Європи близько 40 тис. років тому, зустріли там неандертальців і протягом 15 тисячоліть співіснували з ними до їхнього зникнення в останній льодовиковий період. Нащадки цих кроманьйонців (гаплогрупа I+R) становлять більшу частину населення Європи донині.
Розповсюдження гаплогрупи I.
Розповсюдження гаплогрупи J.
Середньостатистичний араб Мідії Саудівської Аравії. Для гаплогрупи J спостерігається найвищий показник (близько 50%). Національність виникла 20-25 тис. Років тому.

Сенс життя пов'язаний із питанням «Заради чого жити», а не питанням про те, як підтримувати життя. Ставлення людини як...

Гриб - це живий організм, що утворює окреме однойменне царство. Довгий час їх відносили до царства рослин. Але в...

Для любителів «тихого» полювання грибна пора починається раннім літом і триває до пізньої осені. І рідко коли вони...

Олешнікова, В.І. Використання послуг професійних консультантів. - М.: Інфра-М, 1999. - 240 с. 2. Бейч, Е.

Апельсиновий сік. Символічне значення апельсинового соку в сонниках - насолода та спокуса. Досить часто нам...

Різні труднощі, зустрінуті на життєвому шляху, ми найчастіше долаємо. Звичайно, для цього ми додаємо...

Цього року Ваш покровитель Нептун перебуватиме у Вашому сузір'ї і це гарний знак, адже Ви...

1993 кого? 1993 який тварини? — За китайським гороскопом 1873, 1933, 1993 стосувалися років Чорного...

Корпускулярно-хвильовий дуалізм світла означає, що світло одночасно має властивості безперервних...

Роль біології величезна у світі. Хоча вона не входить до пріоритетних предметів, більшість школярів і...
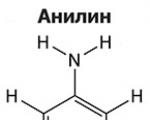
Аміни - органічні похідні аміаку, що містять аміногрупу NH 2 та органічний радикал. У загальному випадку...
Як відповідати на питання частини Друга частина роботи зі суспільствознавства складається з 7 завдань з короткою відповіддю.

Форма ТОРГ-15 складається в тому випадку, коли під час транспортування, переміщення між і всередині складу, при...

Дієтологи кажуть, що для хорошого здоров'я та стрункої фігури, потрібно обов'язково включати перекушування до свого...

Гриб - це живий організм, що утворює окреме однойменне царство. Довгий час їх відносили до царства.

Для любителів «тихого» полювання грибна пора починається раннім літом і триває до пізньої осені. І рідко коли вони...