Composite order Tools and materials
Tuscan order. Tuscan order, one of the five Roman architectural orders. The name is associated with Etruscan architecture...
COMPLEMENT(lat. complementum addition) - a polymolecular system of whey proteins, one of the most important factors of natural immunity. Functions in the blood of humans, cold-blooded and warm-blooded animals. Contained in lymph and tissue fluids. Being included in the composition of immune complexes, K. carries out the lysis of cellular antigens sensitized by antibodies, determines the immune adhesion reaction (see), participates in the opsonization of bacteria, viruses and corpuscular antigens, accelerating their phagocytosis, and participates in the development of inflammation.
K. was first described under the name “alexin” at the end of the 19th century. as a nonspecific thermolabile factor that determines the bactericidal properties of fresh blood serum (G. Bukhner, 1889). The term “complement” was introduced by P. Ehrlich (1900), who believed that the bactericidal factor complements the cytolytic effect of antibodies.
At least 18 proteins are known that make up the K system. These include 9 K components, 8 of which are individual proteins, and one is a complex: 4 proteins of the properdin system, 1 enzyme inhibitor and 2 inactivator enzymes.
According to the nomenclature adopted by WHO, the K system is designated by the symbol C, its individual components by numbers (C1, C2...C9), and fragments of K components by lowercase letters (for example, SZa). The presence of enzymatic activity in a fragment is indicated by a line above its symbol, and the presence of a binding center with the cell membrane is indicated by an asterisk near its symbol [K. F. Austen et al., 1968].
K.'s components circulate in the blood in the form of precursors, without combining with free antibodies or antigens. Two biol, mechanisms of activation (binding) of the K. system are described - classical and so-called. alternative, or properdine [Müller-Eberhard (H. J. Muller-Eberhard), 1975; Vogt (W. Vogt), 1974].
The classical mechanism of K. activation is carried out with the participation of IgG and IgM antibodies that are part of immune complexes, or nonspecifically aggregated immunoglobulins of these classes. When combined with antigens or as a result of nonspecific aggregation, centers that bind C1, the first component of the K system, are formed in the molecules of these immunoglobulins (A. Ya. Kulberg, 1975). Fixed on immunoglobulin C1 initiates a chain of reactions into which the remaining components of the K system sequentially enter.
C1 is a complex of three subcomponents (C1q, C1rr and C1s) formed in the presence of calcium ions; C1q is a collagen-like protein with a mol. weighing (mass) 400,000, consisting of six non-covalently linked identical subunits. Each subunit contains a recognition center for binding to an immunoglobulin molecule. The attachment of C1q to immunoglobulin is accompanied by intramolecular rearrangement of C1q and activation of the associated proenzyme Clr, which acts on C1s proesterase. The resulting C1s-esterase (C1s) affects the second (C2) and fourth (C4) components of K in the liquid phase.
The C4 molecule (molecular weight 208,000) is built from three peptide chains - alpha, beta and gamma, connected by disulfide bonds. C1s cleaves off the C4a peptide from the alpha chain, mol. the weight is 8000, and in the remaining C4b fragment of the molecule a binding center appears with the cell membrane sensitized by antibodies. When C1s acts on C2, mol. the weight of which is 117,000, two fragments are formed - C2b (mol. weight 37,000) and C2a (mol. weight 80,000). In the latter, a binding center for C4b is formed. The C42 complex formed on the cell membrane is capable of cleaving C3; therefore it is called S3 convertase.
The SZ molecule (molecular weight 180,000) is built from two peptide chains - alpha and beta. As a result of the cleavage of C3 peptide from the alpha chain by C3 convertase with mol. weighing 9000 in the C3b fragment of the molecule, a binding center with the cell membrane is formed and a C423 complex with peptidase activity towards C5 (C5 convertase) is formed on the membrane.
After proteolytic cleavage of C5, the assembly of the membrane attack unit begins from the so-called. terminal components of the K system. The C5 molecule is constructed similarly to S3 from two peptide chains a and p, mol. whose weight is 110,000 and 70,000, respectively. C5 convertase cleaves off the C5a peptide from the alpha chain with a mol. weighing 16,500. The resulting C5b fragment has the ability to sorb sequentially one molecule of C6 and C7. The C567 complex sorbs one C8 molecule and six C9 molecules. At the moment of formation, the C5-9 complex attacks the cell membrane, causing its destruction. The cytolytic activity of the complex is determined by C8 and is significantly enhanced by C9.
Along with the cytolytic components, when the K system is activated, the physiologically active peptides C3a and C5a, called anaphylatoxins, are formed; they cause the release of histamine by mast cells. contraction of smooth muscles and increase vascular permeability, and also serve as chemotactic factors for polymorphonuclear cells. Directed migration of polymorphonuclear cells at the site of K. activation is also caused by the trimolecular complex C567 [Ward (P. Ward), 1975]. Another biologically active peptide that appears upon activation of the K system is C3b. When binding to the cell membrane, it acquires a second stable binding center in relation to receptors located on the surface of a number of cells (macrophages, platelets, erythrocytes). This process, called immune adhesion, enhances the phagocytosis of K.-loaded cells and corpuscular particles [S. Ruddy, 1974].
K. also takes part in the mechanism of nonspecific resistance to infections. In this case, the K. system is activated without the participation of antibodies by polysaccharides or lipopolysaccharides that are part of the cell walls of bacteria, yeast, plants, or aggregated IgA. K. binding occurs along an alternative pathway, starting from C3, bypassing the activation stages of C1, C4 and C2. It has been shown that the serum protein properdin, the C3 convertase activator and a number of its precursors take part in the formation of C3 and C5 convertases of the alternative pathway. When K is activated via the alternative pathway, as well as the classical one, the cytolytic complex C5-9 is formed, as well as the physiologically active peptides C3a and C5a. This mechanism probably underlies the nonspecific elimination of viruses and altered erythrocytes from the body [L. Pillemer, 1954, 1955].
All of the indicated functions of the reaction products of the components of K. are aimed at the destruction and rapid removal of inf. from the body. or foreign agents. They determine the significance of the K system as a protective factor of the body.
In addition to its protective function, the K system can contribute to damage to the body’s own tissues in a number of diseases with an autoimmune component (glomerulonephritis, systemic lupus erythematosus, arteritis, myocarditis, endocarditis). In this case, activation of the K. system is carried out both by antibodies directed against tissues and by soluble or fixed immune complexes in tissues. The resulting complexes C423 and C5-9 of the K components are fixed on both sensitized and non-sensitized cells by antibodies, causing destruction of their membranes. An important role in the autoimmune process also belongs to C3a and C5a peptides and the C567 complex [N. R. Cooper, 1974; L. G. Hunsicker, 1974; McCluskey (R. Mc Cluskey), 1975].
The content of K. is most often judged by its hemolytic activity against sheep erythrocytes sensitized with rabbit hemolysin. K.'s titer is expressed in 100 or 50% hemolytic units (CH100 or CH50), i.e., the minimum amount of K., which under the selected standard experimental conditions lyses, respectively, 100 or 50% of optimally sensitized erythrocytes. K.'s content can also be assessed by its cytolytic effect in the lymphocyte - antilymphocyte serum system [Terasaki (R. I. Terasaki), 1964]. K., which does not have lytic activity, for example. K. horse, bull, mouse, can be determined in the agglutination reaction of sensitized erythrocytes loaded with K. with bovine serum protein - conglutinin (see Conglutination).
Individual components of K. are titrated in a hemolytic test using special reagents, which are preparations of fresh guinea pig serum, devoid of only the titrated component, and containing the remaining components in excess. The corresponding hemolysis intermediates can also be used as titration substrates. Immunochemical and titration methods using anti-sera to pure components of K are widely used.
The content of K. in the sera of animals of various species, according to hemolytic titration, varies greatly. Its highest titer, reaching 200 CH50 per 1 ml, was determined in guinea pigs. 1 ml of human serum contains on average 70, and rabbit 20 CH50 [Audran (R. Audran), 1959, 1960]. However, K.'s titers in the hemolytic test do not always correspond to its true content. Thus, K. of some species does not lyse sensitized sheep erythrocytes, although it binds to them. The hemolytic activity of K. of different types is not the same when tested in various hemolytic systems [Boyd (W. S. Boyd), 1969].
Biol, the properties of K. of various types are largely determined by the content of individual components in them. Species differences are especially pronounced in the content of C2 and C4. These components are completely absent or contained in very low titers in the sera of horses, bulls, and mice, which do not have lytic activity. Serums of all types are characterized by a high content of C1. The content of K. components in human serum is determined in weight units.
Individual fluctuations in the level and composition of K. in healthy people aged 8-35 years are insignificant and do not depend on blood type and Rh factor. Typically, women contain 10% less K than men, and in newborns and pregnant women its content is reduced by an average of 30% [J. Gumbreitier et al., 1960, 1961]. There was a tendency to increase K. levels between the ages of 35 and 60 years.
The content of K. in the sera of patients depends on the nature of the disease. In most acute infections of purulent etiology, as well as in staphylococcal bacteremia, an increase in K titers is observed in the initial period. It is believed that this is associated with the activation of cells of the reticuloendothelial system, in particular macrophages that synthesize C2, C4, C5. During the period of elimination of antigens with the participation of antibodies, K.'s titers decrease and reach the norm during recovery. In a number of diseases that affect the cells of the liver parenchyma, for example, cirrhosis, hepatitis, hron, cholecystitis, the synthesis of C3-, C6-, C9- and C1-inhibitors is disrupted, which leads to a decrease in the overall level of K. As a rule, the level of K. decreases with allergic conditions, autoimmune diseases and diseases of immune complexes due to the binding of K. circulating in the blood and associated in tissues with immune complexes. Cases of deficiency in individual components of K., accompanied by various pathol, conditions are described.
The K. system is active in the body and in freshly isolated serums. K. is inactivated within 2-4 days when sera are stored in the refrigerator (t° 5°), and as a result of heating the sera at t° 56° - for 20 minutes. Inactivation of K. under the influence of various physical agents has been described. factors - sunlight, ultraviolet radiation, shaking, under the influence of chemicals. agents - weak solutions of acids, alkalis, organic solvents, proteolytic enzymes (L. S. Reznikova, 1967). K.'s activity remains for a long time in freeze-dried sera, when sodium sulfate (5%) and boric acid (4%) are added to fresh sera, in sera stored at a temperature of -40° and below.
K.'s ability to be included in immune complexes is used to detect antibodies and antigens (see Antigen - antibody reaction, Complement fixation reaction). However, it must be borne in mind that many antisera and some antigens bind K. nonspecifically. This phenomenon, called the anticomplementary effect, is expressed in a decrease in the hemolytic activity of K. It may be due to the admixture of aggregated globulins, lipopolysaccharides or proteolytic enzymes in titrated preparations, as well as bacterial contamination of the preparations (Boyd, 1969). The increased ability of antibodies of some individuals within one species to nonspecific fixation of K. is called deviability, and antibodies that have this property are called deviable.
The study of the activation process of K., clarification of biol, the properties of the activation products of K. components, the level of K. in normal conditions and in various diseases makes it possible to understand its protective function and its role in tissue damage. This knowledge is necessary, in particular, for the development of scientifically based methods for the prevention and treatment of diseases caused by activation of the K system.
Determination of K.'s titers in various diseases over time is of practical importance, since it is an indicator of immunol, the state of the body, the effectiveness of treatment. activities and has prognostic significance.
Bibliography: Boyd W. Fundamentals of immunology, trans. from English, p. 346, M., 1969; Inflammation, immunity and hypersensitivity, ed. G. 3. Moveta, lane. from English, p. 422, M., 1975, bibliogr.; Kulberg A. Ya. Immunoglobulins as biological regulators, p. 106, M., 1975, bibliogr.; CabotE. iMeyer M, Experimental immunochemistry, trans. from English, p. 140, M., 1968, bibliogr.; P e z n and to the island and L. S. Complement and its significance in immunological reactions, M., 1967, bibliogr.; A u s t e n K. F. a. o. Nomenclature of complement, Bull. Wld Hlth Org., v. 39, p. 935, 1968; Col ten H. R. Biosynthesis of complement, Advanc. Immunol., v. 22, p. 67, 1976, bibliogr.; Comprehensive immunology, ed. by N. K. Day a. R. A. Good, v. 2, N.Y., 1977; Muller-Eberhard H. J. Complement, Ann Rev. Biochem., v. 44, p. 697, 1975, bibliogr.; Yogt W. Activation, activities and pharmacologically active products of complement, Pharmacol. Rev., v. 26, p. 125, 1974, bibliogr.
I. A. Tarkhanova.
Structure of the complement system:
1. proteins – activators (initial components) – C1qrs, C2, C3, C4.
2. proteins of the final sequence (terminal components) – C5, C6, C7, C8, C9.
3. factors of the alternative activation pathway – D i, B i, P.
4. proteins inhibitors and activators:
· having enzymatic activity – factor I, C3b – inactivator, carboxypeptidases.
· Binding and competing proteins - C4BP (C4 - binding protein), vitronectin or S - protein, C1 - INH (C1 - inhibitor), factor H, DAF (CD55 - putrefaction-enhancing factor), CD59, MCP (CD46).
C2b is inactivated by C4BP and DAF, C3b is inactivated by CR1 b factor H.
5. receptors for complement proteins. Designated CR - complement receptor.
CR3(CD11b, CD18)
CR4(CD11c, CD18)
The complement system consists of about forty functionally related plasma proteins: components, regulatory protein factors and receptors. They are capable of sequential activation in the form of a cascade with a final nonspecific cytotoxic effect towards bacteria, fungi, viruses, tumor cells, transplant cells and others.
Complement is included in Hagemann megasystems, along with the blood coagulation system, fibrinolytic and kinin - kallikrein system.
Fragments– these are products of the breakdown of components: C3a, C3b, etc. If the fragment is inactivated, then add the letter I: C3ai; proteins with enzymatic activity are indicated by a line above the letter D -, C4α, C4β are individual chains in peptide molecules.
Biosynthesis of complement proteins occurs in the liver, small intestinal epithelium, bone marrow, spleen and macrophages.
Synthesis and consumption, activation and inhibition are in labile equilibrium and are tightly regulated. In some cases, the polypeptide chains of one component C1, C8 are synthesized separately and assembled just before secretion as a binary weapon. Complement synthesis in the embryonic period begins from the 6th week of development, and at the 10th week cytotoxic activity is already recorded.
Mechanisms of complement activation:
1. classical pathway – dependent on immune complexes
2. lectin – close to classical
3. alternative – dependent on factor P
4. protease – close to alternative, but does not depend on factor P.
Classic activation path.
It is triggered by the antigen + antibody complex (immune complex - IR), that is, it requires the presence of a specific immune process at the moment. It can be triggered by the antigen + C-reactive protein complex, or by the mannose-binding lectin (lectin pathway) - MBL. By attaching to the surface of the target cell (*: bacterium), the antibody initially changes what should be subsequently destroyed by complement. The connection of an antibody with an antigen leads to a change in the spatial configuration of the antibody, which makes it possible for further fixation of C1q on it.
However, for strong fixation, 1 IgM molecule or 2 IgG molecules are required. Other classes of Ig do not activate complement.
The C1 complement component consists of three components: C1q, C1r, C1s.
C1q is a monomer, C1r is a dimer (preprotease C1s), C1s is a dimer (preprotease for C2 and C4).
Externally, the C1qrs molecule has the shape of a tulip.
Physiological and pathological effects of activated complement fragments.
C2a is a kinin-like substance that irritates pain receptors.
C4a – causes severe dilation of capillaries and severe tissue swelling
C3b – stimulation of adhesion and opsonization
C3a, C5a are anaphylotoxins 1 and 2, respectively, they stimulate the chemotaxis of cells to the site of inflammation, causing spasm of smooth muscles, increased capillary permeability, and degranulation of mast cells (for example, due to histamine). C5a is a more aggressive fragment than C3a. C5a has the potential for cytolysis (many oxygen-dependent radicals are formed).
C5b6789...9 is a substance in the shape of a hollow cylinder with lipid-binding centers; it is a membrane attack complex. A cylinder with a hole diameter of 10 nm, which, due to lipid-binding centers, is capable of fixing on the membrane of the target cell, drilling it, as a result of which the contents of the cell flow out (more precisely, sodium ions and water enter the cell).
Basic functions of complement.
1. microbicidal – directed against bacteria, viruses, fungi
2. cytolytic - against tumor cells of transplants, in case of pathology - against their own (in case of regulatory defects and insect or reptile bites).
3. participation in inflammation – C2a, C3a, C4a, C5a
4. activation of the phagocytic reaction: chemotaxis, opsonization, adhesion and absorption. The main fragment is C3b, but chemotaxis is also stimulated by C3a and C5a.
5. interaction with other parts of the Hageman megasystem: coagulation factor 12 - capable of activating the complement system through an alternative pathway.
Kallikrein, plasmin, thrombin – activate C3, C4, C5, B.
6. participation in the regulation of the immune response
7. participation in allergic reactions – anaphylotoxins.
Complement is a system consisting of complement proteins themselves, membrane receptors for complement, plasma and membrane regulators of complement activity.
Complement proteins themselves are a series of glycoprotein and protein factors in the blood plasma, including 9 different components. They form a multi-enzyme molecular cascade, in which the product of one reaction is the substrate for the next. In this case, a gradual increase in lytic potential occurs and an initially weak initiating stimulus leads to a powerful final antimicrobial effect.
There are 4 types of receptors for complement components (complement receptor, CR - I, II, III, IV). The first type receptor (CR) is found on the surface of antigen-presenting cells and red blood cells. It mediates the capture of a pathogen to which opsonins C3b and C4b are attached. Binding of immune complexes by erythrocytes ensures their transport to the liver and spleen, where macrophages are contained. The second type receptor (CR II) is expressed on B lymphocytes and follicular dendritic cells. It takes part in the fixation by these cells of immune complexes in the germinal centers of follicles of lymph nodes, causing further somatic hypermutagenesis of immunoglobulin receptors of B-lymphocytes and the formation of memory B-cells. CR III and CR IV by their nature belong to β 2 -integrins (adhesion molecules) and are specific to iC3b (inactivated under the influence of factor H) and C3d. These receptors are found predominantly on phagocytes and perform a dual function. Firstly, they promote the migration of phagocytes to the site of inflammation, since they can interact with the adhesion membrane molecules ICAM-1 and ICAM-2, the expression of which on tissue cells is one of the landmarks for directed movement. Secondly, phagocytes, penetrating into the lesion thanks to CR III CR IV, recognize complement components, which promotes phagocytosis of the opsonin-labeled pathogen.
Along with numerous complement components, proteins with antagonistic properties circulate in the blood plasma, which limit the activation of the complement system during the neutralization of the pathogen. One of the most important is the inhibitor of the first component (C1-inhibitor), the deficiency of which causes an increased risk of developing hereditary angioedema. The so-called factor H ensures the inactivation of C3b, promoting its further cleavage into fragments C3c and C3d, and factor I destroys C3b and C4b.
As you can see, 2 plasma factors neutralize C3b at once. This is necessary for the correct operation of the alternative pathway, since an excess of this fragment causes unreasonable hyperactivation of complement, causing severe self-damage. The initiation of the cascade occurs precisely due to C3b, formed during the spontaneous hydrolysis of C3. It should be noted that spontaneous hydrolysis is always limited, which prevents possible hyperactivation of the system. At the same time, under the action of C3 convertase, the C3b fragment is formed in an amount sufficient to initiate a new cascade, the unfolding of which releases an additional portion of C3b. Due to this positive feedback, complement increases its lytic potential while the amount of pathogen remains unchanged. However, if the described process is not properly controlled, unreasonable hyperactivation of complement along the alternative pathway and, as a result, damage to one’s own tissues is quite possible. Moreover, the alternative mechanism can also be potentiated by C3b, released as a result of the classical activation pathway occurring in parallel. Therefore, for the correct operation of the entire system, adequate inactivation of the resulting C3b is necessary.
Since complement carries out pattern recognition, and the membrane structures of one’s own cells normally undergo dynamic changes, there is a potential danger of complement-mediated auto-aggression. To prevent it, protective proteins are “built in” into the membranes of one’s own cells, inactivating the complement cascade. We are talking about a factor that accelerates decay (English: Decay accelerating factor, DAF), which is found on blood cells, epithelial cells and endothelial cells. It enhances the catabolism of the key enzymes of the cascade - C3 and C5 convertases. Membrane protective proteins also include membrane cofactor protein (MCP), which is a cofactor in the proteolysis of C3b and C4b using factor 1.
The role of the complement system is: Material from the site
Complement system is a complex complex of serum globulins. This cascade system of proteolytic enzymes is intended for the humoral protection of the body from the action of foreign agents and is involved in the implementation of the body's immune response. Complement system proteins provide a quick and effective response to an initially weak signal and bring it to functional consequences. The components of the complement system are usually designated by Latin letters.
There are two mechanisms for activating the complement system:
classical;
alternative.
These mechanisms are connected at the level of the 5th component and then proceed in the same way.
The classic way.
The triggering mechanism is the formation of an antigen-antibody complex (AG-AT) on the surface of the target cell. At the same time, conformational changes occur in the immunoglobulin molecule (it is designated: Ig or AT). As a result of these changes, Ig acquires the ability to bind the C 1 q component of complement. C 1 r and C 1 s join them, and this entire complex undergoes a conformational rearrangement and turns into a C 1 esterase, which acts on C 4, C 4 a is cleaved, and C 4 b is part of the complex. Then C2 joins the complex, forming a new substrate for the action of C1s, C2b is cleaved, and C2a becomes part of the complex.
The resulting complex is called “C 3 -convertase”, and under its action the peptide C 3 a is cleaved, and C 3 b is part of the complex, which is now called “C 5 -convertase”. C5 convertase acts on C5, cleaves C5a from it, and C5b is part of the complex.
After this, C 6, C 7 and C 8 are sequentially associated with C 5 b. As a result, a complex is formed that is capable of attaching 2 C9 molecules.
If this process occurs on the surface of the target cell, then the components of the C 5 b-C 9 complex form a membrane attack complex, which forms transmembrane channels on the surface of the target cell that are completely permeable to electrolytes and water. The target cell dies.
By-products (minor) of the process C 3 a and C 5 a have the properties of anaphylotoxins.
Regulation of the classical pathway.
Most components are active only as part of the complex. Their active forms can exist for a very short time. If during this time they do not meet the next component, then the active forms lose contact with the complex and become inactive. If the concentration of any component is below the threshold (critical), then the operation of the complement system will not lead to physiological consequences.
Endogenous proteinase inhibitors also take part in the regulation of the complement system. The most effective of them is the C 1 inhibitor.
Alternative way.
The difference between the alternative pathway and the classical one is that it does not require the formation of immune complexes to trigger it.
The triggering mechanism of the alternative pathway is the formation of C 3 b from C 3 under the influence of some trigger factor: for example, polysaccharides of the bacterial cell wall.
C3b forms a complex with factor “B” (C 3 bB), which is exposed to protease D (always active in blood plasma!). As a result, “Ba” is cleaved off and the C3bBb complex is formed, which has proteolytic activity towards C5 - it cleaves C5a from it.
After this, the reactions proceed in the same way as in the classical way.
The substrate for C 3 b is also C 3, as a result of which an even larger amount of C 3 b is formed - a positive feedback is observed. Therefore, even small amounts of C 3 bBb are sufficient to obtain more and more of its active form (amplification of the initially weak signal).
The alternative pathway normally works always and very actively, which provides a rapid nonspecific response to the introduction of foreign cells.
Specific inhibitors take part in the regulation of the complement system, which regulate the speed of enzymes of key reactions.
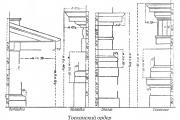
Tuscan order. Tuscan order, one of the five Roman architectural orders. The name is associated with Etruscan architecture...

LIBRETTO. If you love music (which you undoubtedly do, since you needed this book), then you've probably had...

Religion: paganism Birth: 942 (0942) Death: March on the Dnieper Family: Rurikovich Father: Igor...

Masaccio (actually Tommaso di Giovanni di Simone Cassai (Guidi), Tommaso di ser Giovanni di Guidi; December 21, 1401,...

(Published in Tel Aviv in 1958. Transmitted with minor abbreviations). Jews! Love each other, help each other...

Take a ladle or saucepan and pour milk and cream into it. Add vanilla sugar (sugar with vanilla seeds). In a separate...

Hello everyone! Today we have a very tasty and simple recipe in front of us - sausages in dough in the oven. Do you feel...

Avocado is one of the healthiest fruits. This is a real storehouse of vitamins and nutrients. And him...

Roast pork is an ideal tasty and satisfying dish that can feed the whole family. Mistresses...

This cake is very easy to make and tastes like the classic Napoleon. When preparing the cake there is...

Representatives of these zodiac signs do not always become close people, but they are pleasant and interesting to each other....

Compatibility between a Rooster man and a Pig woman is based on mutual understanding between the partners. They have different personalities and...

several basic concepts and formulas. All substances have different mass, density and volume. A piece of metal...

If we’re going to talk about gender equality, then it’s with someone like an Aries woman. This emancipated lady is a bright...

LIBRETTO. If you love music (which you undoubtedly do, since you needed this book), then you'll probably...

Religion: paganism Birth: 942 (0942) Death: March on the Dnieper Family: Rurikovich Father:...