Filosofinė problemos vizija
Gyvenimo prasmė susijusi su klausimu „Dėl ko gyventi“, o ne su klausimu, kaip išlaikyti gyvybę. Žmogaus požiūris...
Rašyti šį straipsnį mane paskatino nenutrūkstantys pokalbiai, kad ukrainiečiai yra slavai, o rusai visai ne slavai, o jau seniai mongolai.
Natūralu, kad tokių ginčų iniciatoriai yra vadinamieji Ukrainos patriotai. Šiuo atveju išvados daromos remiantis kai kurių naujai nukaldintų istorikų teorijomis, iki šiol nežinomais istoriniais dokumentais ir kt. Bet be istorijos, o dažnai ir pseudoistorijos, yra ir toks mokslas kaip genetika, su genetika nesiginčysi, mielieji.Taigi, norime to ar nenorime, turime tą patį genotipą.
Kas yra haplogrupė?
Y-chromosomų haplogrupės, išpopuliarėjusios biopolitiniuose sluoksniuose, yra statistiniai žymenys, padedantys suprasti žmonių populiacijų kilmę. Tačiau daugeliu atvejų toks žymuo nieko nesako apie individo etninę kilmę ar rasę (skirtingai nuo kitų DNR analizės metodų). Konkrečios haplogrupės nešiotojų visumoje įžvelgti etninę kilmę, subetniškumą, rasę ar kitą panašaus pobūdžio vienybę ir šiuo pagrindu bandyti sujungti kažkokią tapatybę yra nesąmonė. Ir, žinoma, haplogrupė jokiu būdu „neatspindi asmens dvasioje“.
Y chromosomos ypatumas yra tas, kad ji perduodama iš tėvo sūnui beveik nepakitusi ir nėra "sumaišyta" ar "atskiesta" dėl motinos paveldimumo. Tai leidžia jį naudoti kaip matematiškai tikslų įrankį tėvo kilmei nustatyti. Jei terminas „dinastija“ turi kokią nors biologinę reikšmę, tai būtent Y chromosomos paveldėjimas. (Norėdami gauti išsamų, bet lengvai suprantamą reiškinio paaiškinimą, spustelėkite nuorodą)
Y chromosoma yra kitas dalykas: ji susideda iš genų, tiesiogiai atsakingų už vyrų reprodukcinę sistemą, o menkiausias defektas, kaip taisyklė, daro vyrą sterilų. „Santuoka“ toliau neperduodama, o Y chromosoma „išsivalo“ kiekvienoje kartoje.
Tačiau be žalingų mutacijų, vyriškoje chromosomoje kartas nuo karto atsiranda neutralių mutacijų, į kurias neatsižvelgiama natūralios atrankos. Jie yra susitelkę „šiukšlių“ chromosomos regionuose, kurie nėra genai. Kai kurios iš šių mutacijų, įvykusių prieš 50–10 tūkstančių metų, pasirodė esąs patogūs žymenys, leidžiantys nustatyti senovės protėvių populiacijas, kurios vėliau išplito visoje Žemėje ir suformavo šiuolaikinę žmoniją.
Y-chromosomų haplogrupė nustato vyrų rinkinį, kurį vienija tokio žymens buvimas, t.y. kilęs iš bendro patriarchalinio protėvio, kuris prieš daugelį tūkstančių metų turėjo specifinę Y chromosomos mutaciją.
http://en.wikipedia.org/wiki/Human_Y-chromosome_DNA_haplogroup
HAPLOGRUPĖS R1a1 KILMĖ – PIETŲ RUSIJA!
Bet kuri šiuolaikinė etninė grupė susideda iš kelių, mažiausiai dviejų ar trijų Y-chromosomų haplogrupių atstovų.
http://en.wikipedia.org/wiki/Y-DNA_haplogroups_by_ethnic_group
Geografinis haplogrupių pasiskirstymas yra susijęs su senovės populiacijų, kurios tapo protėviais etninėms grupėms ar etninių grupių grupėms, migracijos istorija. Pavyzdžiui, haplogrupė N3 gali būti vadinama „finougrikais“: jei ji randama tarp tam tikros srities atstovų, tai reiškia, kad anksčiau ten gyventojai maišėsi su finougrų tautomis. O gal čia atėjo „mišrios“ gentys.
Haplogrupių statistikos tyrimas leido antropologams atkurti žmonių populiacijų migracijos per pastaruosius dešimtis tūkstančių metų vaizdą, pradedant nuo Afrikos protėvių namų. Tačiau duomenys taip pat gali būti naudojami įvairiems rasistiniams ir ksenofobiniams mitams paneigti.
R1a haplogrupės etnogeografinis pasiskirstymas
Šiuo metu dideli R1a haplogrupės dažniai aptinkami Lenkijoje (56 proc. gyventojų), Ukrainoje (50–65 proc.), Europos Rusijoje (45–65 proc.), Baltarusijoje (45 proc.), Slovakijoje (40 proc.), Latvijoje ( 40 proc., Lietuva (38 proc.), Čekija (34 proc.), Vengrija (32 proc.), Kroatija (29 proc.), Norvegija (28 proc.), Austrija (26 proc.), Švedija (24 proc.), Šiaurės rytų Vokietija ( 23 proc. ir Rumunijoje (22 proc.).
Labiausiai paplitęs Rytų Europoje: tarp luzatų (63 proc.), lenkų (apie 56 proc.), ukrainiečių (apie 54 proc.), baltarusių (52 proc.), rusų (48 proc.), totorių 34 proc., baškirų (26 proc.). %) (tarp Saratovo ir Samaros regionų baškirų iki 48%); ir Vidurinėje Azijoje: tarp chudžando tadžikų (64 proc.), kirgizų (63 proc.), iškašimų (68 proc.).
Halogrupė R1a būdingiausia slavams. Pavyzdžiui, tarp rusų yra paplitusios šios haplogrupės:
R1a – 51 % (slavai – arijai, lenkai, rusai, baltarusiai, ukrainiečiai)
N3 – 22 % (suomiai-ugrai, suomiai, balti)
I1b – 12 % (normanai – vokiečiai)
R1b – 7 % (keltai ir kursyvas)
11a - 5% (taip pat skandinavai)
E3b1 – 3 % (Viduržemio jūros regionai)
Labiausiai paplitusi haplogrupė tarp ukrainiečių:
R1a1 - apie 54% (slavai - arijai, lenkai, rusai, baltarusiai, ukrainiečiai)
I2a – 16,1 % (balkanų tautos, frakai, ilirai, rumunai, albanai, graikai)
N3 – 7 % (suomiai-ugrai)
E1b1b1 – 6 % (Afrikos tautos, egiptiečiai, berberai, kušnyrai)
N1c1 – 6 % (Sibiro tautos, jakutai, buriatai, čiukčiai)
Kaip rodo tyrimai, pagal Y chromosomos žymenis tirti ukrainiečiai genetiškai labiausiai artimi kaimyniniams pietvakarių rusams, baltarusiams ir rytų lenkams. Trys slaviškai kalbančios tautos (ukrainiečiai, lenkai ir rusai) sudaro atskirą klasterį pagal Y haplogrupes, o tai rodo bendrą išvardytų etninių grupių kilmę.
Mitai.
Visi žino mitą, kad rusai daugiausia yra mongolų, senovėje pavergusių Rusiją, palikuonys. Haplogrupių statistika nepalieka akmenų šiam mitui, nes tipiškų „mongoloidinių“ haplogrupių C ir Q tarp rusų apskritai nėra. Tai reiškia, kad jei mongolų kariai kažkada atvyko į Rusiją su antskrydžiais, tai visos jų sugautos moterys buvo nužudytos arba išvežtos kartu su jomis (kaip ir Krymo totoriai vėlesniais laikais).
Kitas paplitęs mitas, kad rusai Centrinėje ir Šiaurės Rusijoje didžiąja dalimi yra ne slavai, o suomių-ugrų aborigenų palikuonys, kurių jūroje tariamai dingo keli slavai. Iš čia jie kildina „rusišką girtumą“, „rusišką tinginystę“ ir kt. Tuo tarpu „suomiškos“ haplogrupės N3 dalis tarp rusų Centrinėje Rusijoje siekia apie 16% (retai apgyvendintuose regionuose į šiaurę nuo Maskvos vietomis siekia 35%, o tankiai apgyvendintuose regionuose į pietus ir vakarus nuo Riazanės sumažėja iki 10). %). Tie. iš šešių tėvų tik vienas buvo suomis. Galima daryti prielaidą, kad santykis motinos genofonde yra maždaug toks pat, nes slavai ir finougrai, kaip taisyklė, taikiai sugyveno.
Beje, Tarp Suomijos suomių haplogrupė N3 yra atstovaujama maždaug 60% gyventojų.. Tai reiškia, kad iš penkių tėvų du buvo ne „originalūs suomiai“, o „praeinantys bičiuliai“, galbūt duoklės rinkėjai iš Novgorodo. Tarp etninių estų ir latvių „suomių tėvų dalis“ dar mažesnė – apie 40 proc. Vokiečių ir slavų kilmės „praeinantys bičiuliai“ aiškiai dominavo karštuose estų vaikinuose. Bet lietuvaitės jas pamilo: lietuvės, nepaisant indoeuropiečių kalbos, yra tie patys 40% finougrų palikuonys.

Tarp etninių ukrainiečių taip pat yra „suomių tėvų dalis“, nors tris kartus mažesnė nei tarp rusų. Tačiau suomių-ugrų gentys Ukrainoje negyveno, o ši dalis buvo atvežta iš Vidurio Rusijos. Bet jei „suomiško kraujo dalis“ tarp etninių ukrainiečių yra tik tris kartus mažesnė nei tarp rusų, tai mažiausiai trečdalis jų yra rusų tėvų palikuonys. Matyt, anksčiau „neatsakingos“ pietų rusų merginos mėgo kvailioti su „maskvėnų okupantais“. Kol ukrainiečių berniukai linksminosi Zaporožės sichoje vyrų kompanijoje, jų seserys ir dukros suprato draugiškus Suvorovo stebuklų herojus, turinčius sunkias suomių Y chromosomas.
Padėdamas suprasti tam tikrų mitų nenuoseklumą, haplogrupės savo ruožtu gali paskatinti naujų mitų kūrimą. Yra žmonių, kurie jiems suteikia rasinę reikšmę. Svarbu suprasti, kad pačios haplogrupės negali būti rasinės, etninės ar subetninės tapatybės kriterijus. Pritaikius konkrečiam žmogui, jie visiškai nieko nesako. Pavyzdžiui, negali būti suformuota tinkama bendruomenė, vienijanti žmones iš „arijų“ haplogrupės R1a1. Ir atvirkščiai, nėra jokio objektyvaus skirtumo tarp tame pačiame regione gyvenančių rusų, „suomiškos“ haplogrupės N nešiotojų, ir rusų, „arijų“ haplogrupės R1a nešiotojų. Visas kitas „protėvių suomių vyrų“ ir „protėvių arijų vyrų“ palikuonių genofondas jau seniai buvo sumaišytas.
Iš daugiau nei 20 000 žmogaus genomo genų tik apie 100 yra įtraukti į Y chromosomą. Jie daugiausia koduoja vyrų lytinių organų struktūrą ir funkcionavimą. Kitos informacijos ten nėra. Veido bruožai, odos spalva, psichikos ir mąstymo ypatumai registruojami kitose chromosomose, kurios paveldėjimo metu pereina per rekombinaciją (tėvo ir motinos chromosomų dalys susimaišo atsitiktinai).
Jei tam tikros etninės grupės atstovai priklauso kelioms haplogrupėms, tai nereiškia, kad ši etninė grupė yra mechaninis skirtingų genofondų populiacijų derinys. Likusi jų genofondo dalis, išskyrus Y chromosomas, bus sumaišyta. Subtilūs skirtumai tarp skirtingų Rusijos haplogrupių atstovų gali būti įdomūs tik tiems, kurie profesionaliai specializuojasi pūtimo srityje.
Ir atvirkščiai, žmonės iš tos pačios haplogrupės gali priklausyti skirtingoms etninėms grupėms ir net skirtingoms rasėms, o jų genotipas ir fenotipas turi esminių skirtumų.
Pavyzdžiui, „arijų“ haplogrupės buvimo rekordininkai yra tokios nepanašios tautos kaip lenkai (56,4%) ir kirgizai (63,5%). „Arijų“ haplogrupė randama daugiau nei 12% aškenazių žydų, ir ne kai kuriuose „puskraujiuose“, o tikriausiuose, tipiškiausiuose jų etninės grupės atstovuose.
Jei rusų jūreivis, apsilankęs Angoloje, gimtinei moteriai „padovanoja“ berniuką, tada jis ir visi jo palikuonys iš vyriškos giminės turės tėvo haplogrupę. Pasikeis 1000 kartų, palikuonys visais atžvilgiais virs tipiškiausiais angoliečiais, bet vis tiek nešios „arijų“ Y chromosomą. Ir šio fakto negalima atskleisti niekaip kitaip, išskyrus DNR analizę.
Tolimoje praeityje haplogrupės R1a1 nešiotojai, šiuolaikinių indoeuropiečių protėviai, keliavo iš Pietų Rusijos ir Uralo tyrinėti Europos, Vidurinių Rytų, Irano, Afganistano, Indijos ir kitų kaimyninių šalių, kurių gyventojams jie primetė savo gyventojus. papročius ir perdavė savo kalbą. Bet jei jų istorinė sėkmė buvo kažkaip susijusi su pažangia biologija (tarkime), tada ji buvo įsišaknijusi ne Y chromosomos ypatybėse, o kituose genuose, kurie buvo protėvių populiacijoje. Šis „išplėstinis genofondas“ buvo susijęs su tam tikra haplogrupe tik statistiškai. Šiuolaikiniams R1a1 haplogrupės atstovams gali trūkti šių „pažangių“ genų. „Arijų“ chromosomos turėjimas jokiu būdu neatspindi „dvasios“.
Tos Y chromosomos sekcijos, kurios tarnauja kaip žymenys identifikuojant haplogrupes, pačios nieko nekoduoja ir neturi jokios biologinės reikšmės. Tai yra gryniausios formos žymekliai. Juos galima palyginti su oranžiniais ir žaliais šviesos diodais filme „Kin-Dza-Dza“, kuriais buvo atpažįstami chaatlanai ir patsakai, ir tarp šių „rasių“ nebuvo jokio kito skirtumo, išskyrus lemputės spalvą. Taigi „arijų“ haplogrupės buvimas pats savaime negarantuoja žmogui ne tik arijų smegenų, bet net ir arijų penio („rasistiniai blowjobs“ gali nusivilti).
Žmogaus DNR yra 46 chromosomos, kiekvienas žmogus pusę chromosomų paveldi iš tėvo, pusę iš motinos. Iš 23 chromosomų, gautų iš tėvo, tik vienoje – vyriškoje Y chromosomoje – yra nukleotidų rinkinys, kuris perduodama iš kartos į kartą be pokyčių tūkstančius metų.
Genetikai DNR rinkinį, kuris paveldimas be pakitimų, vadina haplogrupe.
DNR tyrimai sujungė visus žmones Žemėje į genealogines grupes ir pažymėjo juos raidėmis. Tos pačios haplogrupės žmonės turi vieną bendrą protėvį tolimoje priešistorėje.
Haplogrupė dėl savo paveldimo nekintamumo yra vienoda visiems tų pačių žmonių vyrams.
Kiekviena biologiškai savita tauta turi savo haplogrupę, kuri skiriasi nuo kitų tautų haplogrupių. Haplogrupė yra visos tautos genetinis žymeklis.


Čia yra VISŲ tipų Y-DNR haplogrupių sąrašas, kurie atsirado tam tikrose teritorijose ir po kurio laiko išplito visoje Europoje ir Azijoje.
Haplotipas:
KAM– atsirado prieš 40 000 metų Šiaurės Irane.
T- atsirado prieš 30 000 metų Raudonosios jūros pakrantėse ir (arba) aplink Persijos įlanką.
J– atsirado prieš 30 000 metų Artimuosiuose Rytuose.
R– atsirado prieš 28 000 metų Vidurinėje Azijoje.
E1b1b– atsirado prieš 26 000 metų Šiaurės Rytų Afrikoje.
aš- atsirado prieš 25 000 metų Balkanuose.
J1 - atsirado prieš 20 000 metų Tavros/Zagros kalnuose.
J2– atsirado prieš 19 000 metų šiaurinėje Mesopotamijoje.
E-M78– atsirado prieš 18 000 metų šiaurės rytų Afrikoje.
R1b- atsirado prieš 18 000 metų palei pakrantę Kaspijos jūra ir (arba) Centrinėje Azijoje.
R1a– atsirado prieš 17 000 metų Rusijos pietuose.
G– atsirado prieš 17 000 metų Artimuosiuose Rytuose.
I2– atsirado prieš 17 000 metų Balkanuose.
E-V13– atsirado prieš 15 000 metų pietų Libane arba Šiaurės Afrikoje.
I2b - atsirado prieš 13 000 metų Vidurio Europoje.
N1c1 — atsirado prieš 12 000 metų Sibire.
E-M81– atsirado prieš 11 000 metų Šiaurės Vakarų Afrikoje.
I2a– atsirado prieš 11 000 metų Balkanuose.
G2a– atsirado prieš 11 000 metų Libane arba Anatolijoje.
R1b1b2– atsirado prieš 10 000 metų Kaukazo šiaurėje arba pietuose.
I2b1– atsirado prieš 9000 metų Vokietijoje.
I2a1– atsirado prieš 8000 metų Pietvakarių Europoje.
I2a2– atsirado prieš 7500 metų Pietryčių Europoje.
I1– atsirado prieš 5000 metų Skandinavijoje.
R1b-L21– atsirado prieš 4000 metų Vidurio ir Rytų Europoje.
R1b-S28– atsirado prieš 3500 metų Alpių regione.
R1b-S21- atsirado prieš 3000 metų Fryzijoje (Fryzijoje) Šiaurės jūros pakrantėje ir Vidurio Europoje.
I2b1a– atsirado prieš mažiau nei 3000 metų Didžiojoje Britanijoje.
Europos genofondo analizę pradedame nagrinėdami atskirų haplogrupių genogeografiją. Tai iš dalies yra ankstesnio skyriaus, kuriame trumpai apibendrinta tai, kas buvo žinoma apie Y-chromosomų haplogrupių pasiskirstymą prieš mūsų tyrimą, tęsinys, tačiau dabar pažvelgsime į modernius, išsamius visų pagrindinių Europos haplogrupių ir tų subvariantų pasiskirstymo žemėlapius. kurių dažniai jau žinomi daugelyje Europos gyventojų.
Mūsų Y-bazės duomenų bazėje yra beveik visa paskelbta informacija apie Y-chromosomų haplogrupių dažnius pasaulyje (o ypač Europoje), tačiau ne visa ši informacija tinka geografiniam haplogrupių pasiskirstymui tirti. Kaip ir kuriant kitus genogeografinius atlasus, siekiant sukurti Y chromosomos pasiskirstymo Europoje žemėlapius, duomenys buvo kruopščiai atrinkti ir sukurti. duomenų masyvas žemėlapiams Y chromosomos Europoje. Panagrinėkime kriterijus, pagal kuriuos buvo suformuotas šis duomenų masyvas.
DUOMENŲ RINKINYS ŽEMĖLAPIMS: GYVENTOJAI.
Norint patikimai nustatyti unparentalinių genetinių žymenų (Y-chromosomos ir mtDNR) dažnį, reikalingas dvigubai didesnis mėginio tūris nei autosominiams žymenims – todėl apatinė mėginio riba turėtų būti ne mažesnė kaip 70-100 mėginių. Maži mėginiai buvo sujungti su geografiškai gretimais tų pačių žmonių pavyzdžiais arba (jei derinti nebuvo įmanoma) nebuvo įtraukti į analizę. Dėl to vidutinis imties dydis žemėlapio duomenų rinkinyje yra N=136 mėginiai. Tačiau buvo padarytos taisyklės išimtys. Kai kurioms tautoms ar regioninėms tautos grupėms, kurias, siekiant išsamumo, reikėjo pateikti žemėlapyje, buvo prieinami tik nedideli pavyzdžiai. Tada, nenoriai, į žemėlapių masyvą turėjome įtraukti mažesnius pavyzdžius. Tačiau tokios išimtys buvo daromos retai – iš 251 kartografuotos populiacijos tik dešimtadalio (29 populiacijų) imties dydis yra mažesnis nei N = 50, o didžiosios daugumos (192 imčių) imties dydis yra N = 70 ir didesnis.
Jeigu apie tautą buvo duomenų apie jos regionines populiacijas, tai kartografavimui šios tautos, kurios geografinė padėtis nežinoma, imtys nebuvo naudojamos (jei publikuodami autoriai tik nurodė savo priklausomybę tautai).
Dėl to į žemėlapių masyvą buvo įtraukti duomenys apie 251 populiaciją (imtį), o bendras mėginių skaičius visose imtyse buvo 34 294 mėginiai. Verta paminėti, kad nemaža dalis šių duomenų buvo gauta mūsų komandos, vadovaujamos šių eilučių autoriaus, tyrimu: 45 populiacijos ir 4880 mėginių. Tai yra mūsų duomenys apie Rusijos populiacijas, ukrainiečių, baltarusių ir kai kurias papildomas Rusijos populiacijas, Šiaurės Kaukazo populiacijas, taip pat mūsų dar neskelbti duomenys apie Krymo ir Kazanės totorių, mokšų ir erzų populiacijas. (Duomenys apie haplogrupių dažnius iš trijų publikuotų straipsnių pateikti 2.1, 2.2, 2.3 lentelėse.) Pasirodo, kad iš viso duomenų apie Y chromosomų kintamumą Europoje masyvo kas penktą populiaciją ir kas septintą mėginį tyrė mūsų komanda (2.1 pav. „mūsų » populiacijos pavaizduotos mėlynais apskritimais), nors kelios dešimtys laboratorijų visoje Europoje vykdė ir vykdo šios srities tyrimus. Jei pridėsime, kad Europa yra labiausiai ištirtas pasaulio regionas, tai iš dalies paaiškina, kodėl mūsų komanda ėmėsi bendros Europos genofondo analizės. Žemėlapiams sudaryti taip pat buvo naudojami Y-bazės duomenys ne tik apie Europos populiacijas, bet ir apie šalia esančių regionų populiacijas, įtrauktas į žemėlapio plotą.
Be 251 Europos populiacijos, tirtos naudojant haplogrupių grupes, apimančias visą jų spektrą, dar apie 200 populiacijų buvo ištirta naudojant tik vienos haplogrupės subvariantus. Tai daugiausia straipsniai, skirti haplogrupėms R1b ir R1a, ir nemažai straipsnių apie haplogrupes N, E ir J. Paprastai pavyzdžiai, naudojami šiuose straipsniuose nuodugniam atskirų haplogrupių genotipui nustatyti, anksčiau buvo tiriami visame pasaulyje. haplogrupių spektras. 2.1 pav. suteikia supratimą apie Europos ir gretimų regionų aprėpties tankį populiacijomis, tirtomis ne pagal vieną haplogrupę, o per visą jų spektrą su įprastu filogenetinės skiriamosios gebos lygiu.
ŽEMĖLAPIŲ DUOMENŲ MASYS: HAPLOGRUPĖS.
Ši haplogrupių tyrimo „įprasto filogenetinio lygio“ samprata yra labai sąlyginė ir kiekvienais metais šis lygis auga. Jei 2000 m. tipiniame straipsnyje haplogrupių skaičius nesiekė keliolikos, tai 2010-aisiais daugumoje straipsnių naudojamos 40–70 haplogrupių plokštės. Žinoma, maždaug nuo 2013 m. žinomų haplogrupių, aptiktų atlikus visą seką, skaičius jau siekia tūkstančius. Tačiau jų atradimas yra tik pirmasis žingsnis, kuriam reikia daug tolesnių žingsnių ir pastangų, kad būtų galima išanalizuoti daugybę labai skirtingų populiacijų, kad būtų galima rasti naujų haplogrupių. Ir kol jų dažnis nebuvo nustatytas įvairiose populiacijose, šių naujai atrastų haplogrupių reikšmė genofondo tyrimams yra nedidelė.
Vienoje lentelėje sujungus dešimčių skirtingų metų straipsnių duomenis, neišvengiamai paaiškėja, kad kai kurios haplogrupės buvo ištirtos labai didelėje populiacijų dalyje, o kai kurios – tik kelioms populiacijoms. Laimei, tai nekelia didelių problemų kartografuojant (beveik visos kartografavimui atrinktos haplogrupės buvo ištirtos dviejose ar trijose dešimtyse populiacijų ir tokio atskaitos taškų skaičiaus paprastai pakanka pagrindinėms tendencijoms nustatyti). Tačiau vis dėlto, svarstant žemėlapius, verta turėti omenyje, kad skirtingų haplogrupių šaltinio duomenų detalės nėra vienodos.
Kartografavimui buvo atrinktos haplogrupės, kurios sudarė pastebimą genofondo dalį bent kai kuriose Europos populiacijose ir buvo tiriamos, kaip ką tik minėta, keliose dešimtyse populiacijų. Šiuos kriterijus atitiko 40 haplogrupių: E1b1a1-DYS271, E1b1b1a1b-L142.1, E1b1b1a1c-V22, E1b1b1b1-M81, E1b1b1c-M123, G2a1-P16, G2a61-L3a, I2a31-P0 a-P37.2, I2b-L35, J1-L255, J1c3-P58, J2-L228, J2a-L152, J2a3b-M67, J2a3h-L207.1, J2b-L282, L-M11, N1b-P43, N1c-M412, O3 Q-M242, R1a1-L120, R1a1a1g-M458, R1a-M558, R1a-Z284, R1a-Z93, R1b1a1-M478, R1b1a2-L265, R1b-L10, R1b1a-M5a1a7, R1b1a-2a1a1a1a 1b1a2a1a1b-P312, R1b1a2a1a1b2a - M153, R1b1a2a1a1b2b1-M167, R1b1a2a1a1b4b-M222, R1b1c-V88, R2a-L266, T-L206.
2.1 lentelė. Mūsų duomenys apie Y-chromosomos haplogrupių dažnį Rusijos populiacijose
2.2 lentelė. Mūsų duomenys apie Y-chromosomų haplogrupių dažnį baltų-slavų tautų populiacijose (paskelbta m.).

2.3 lentelė. Mūsų duomenys apie Y-chromosomos haplogrupių dažnį Kaukazo populiacijose


Ryžiai. 2.1. Populiacijų geografinė padėtis Europoje ir gretimuose regionuose tirta Y chromosomos polimorfizmu. Mėlyni apskritimai rodo mūsų pačių duomenis, žali apskritimai rodo duomenis iš literatūros.
HAPLOGRUPĖR1 a(neskirstant į pošakius).
Kaip nurodyta ankstesnių darbų apžvalgoje, haplogrupė R1 a yra viena iš dviejų labiausiai paplitusių haplogrupių Europoje. Be to, tai yra labiausiai paplitusi haplogrupė visoje Eurazijoje. Jo genotipui nustatyti naudojami skirtingi žymenys – M198, M17, SRY1542, tarp kurių filogenetinio skirtumo beveik nėra, ir yra žinoma daug kitų žymenų, apibrėžiančių šią haplogrupę, todėl bet kuris iš jų gali būti naudojamas jai žymėti. Žemėlapių duomenų masyve jis atrodo kaip R1 a- L120 .

Fig. 2.2 matote aiškią geografinio pasiskirstymo tendenciją R1 a- L120 : Nors jis paplitęs visoje Europoje, platus maksimalaus dažnio plotas yra Rytų Europoje, o šiose populiacijose jis sudaro beveik pusę genofondo. Rytuose maksimalią zoną riboja Volga, o per visą jos ilgį – nuo Aukštutinės Volgos iki žemupio. Pietuose maksimali zona siekia beveik Juodąją jūrą, šiaurės vakaruose – iki Baltijos jūros, o pietvakariuose apsiriboja Karpatais ir Alpėmis. Tačiau įdomiausios šios srities ribos yra vakaruose: nors ten nėra geografinių kliūčių, aukšto dažnio zona R1a vis dar gana aiškiai apribota, o dažnio kritimas maždaug du šimtus-tris šimtus kilometrų siekia beveik 30% ( nuo maždaug 35-45% lenkams ir sorbams iki 10-15% tarp vokiečių). Keli straipsniai buvo skirti šios „Lenkijos ir Vokietijos genetinės sienos“ analizei.
Maksimalių dažnių zonos R1a ribos šiaurėje dar nepaminėjau - nors dažnių skirtumas ten nėra toks ryškus kaip vakaruose (ne nuo raudonos iki žalios, o tik nuo raudonos iki geltonos spalvos tonų, t.y. penkiolika procentų), tačiau atsiskleidžia ir šiaurinė siena Žemėlapis gana aiškus. Šiuo atveju jis praeina vienos tautos – rusų – teritorijoje, pažymėdamas skirtumus tarp Rusijos Šiaurės ir kitų Rusijos populiacijų genofondo. Šie geltoni vidutinio dažnio tonai užima ne tik Rusijos šiaurę, bet ir visą Rytų Europos erdvę tarp Volgos ir Uralo ir tik už Uralo dažnių diapazono. R1 a nukrenta iki tų pačių žemų verčių (žali tonai žemėlapyje), kaip ir Vakarų Europoje, Skandinavijoje, Balkanuose ir Vakarų Azijoje.
HAPLOGRUPĖS SUBVARIANTAIR1 a
Tokiame didžiuliame haplogrupių asortimente R1a-L120 Neabejotinai yra beveik tiek pat šios haplogrupės subvariantų, kurių daugelis gali turėti skirtingus ir santykinai mažus geografinius diapazonus. Tačiau laikui bėgant potipiai viduje R1 a buvo nežinomi. Kol Peterio Underhillo laboratorijos Stanfordo universitete darbo dėka iš pradžių buvo atrasti pirmieji du, o paskui dar keli subvariantai. Žinoma, šiuo metu žinomų subvariantų skaičius R1 a skaičiai šimtais: kiekvienas mėginys, kurio visas genomas arba Y chromosoma buvo visiškai sekvenuota, filogenetiniame medyje sudaro savo šaką. Tačiau kol bent kai kurie iš šių variantų nebuvo ištirti dėl jų paplitimo populiacijose ir nenustatyti jų atsiradimo skirtingose Europos populiacijose dažniai, genogeografinių žemėlapių sudarymas neįmanomas. Dabar galime tik spėlioti apie šių pošakių paplitimo sritis, remdamiesi kartais netvirta informacija apie tų pavienių mėginių, iš kurių buvo atrasta kiekviena iš šių šimtų šakų, kilmės vietas. Susidomėjimas šia haplogrupe yra toks didelis, kad, žinoma, tokia populiacijos patikra artimiausiais metais bus atliekama įvairiose pasaulio laboratorijose (įskaitant mūsų komandą), o vėliau – subvariantų genogeografija. R1 a bus daug detalesnis. Tačiau šiuo metu apsiribojame tais žymenimis, dėl kurių buvo atlikta masinė gyventojų patikra straipsniuose ir mūsų komandos darbe: tai trys pagrindiniai Europos šakos žymenys. R1 a ir Azijos šakos žymeklis.
AZIJOS FILIALASR1 A- Z93.
Reikėtų pažymėti, kad filogenetiniame medyje atsiranda padalijimas į Europos ir Azijos šakas R1 a labai aiškiai. Azijos šakos žymeklis yra Z93 (ir jo sinonimai), o Europoje šios šakos beveik nėra. Norėdami tai įrodyti, pateikiame jo pasiskirstymo žemėlapį (2.3 pav.), kuriame rodomi tik žemi dažniai (1-3%) ir tik labai keliose Rytų Europos populiacijose, o pereiname prie Europos subvariantų svarstymo.

Ryžiai. 2.3. Haplogrupių geografijasR- Z93 Y chromosomos vidujeEuropa.
EUROPOS SKYRIUSR1 A-M458.
Pirmasis, R-M458, identifikuoja Vidurio Europos subvariantą bendrame haplogrupės masyve R1a, užimantis pusę Europos. Todėl, atradę šį žymenį, daugelyje slavų ir Šiaurės Kaukazo populiacijų jį specialiai genotipavome, todėl pirmą kartą buvo galima sudaryti išsamų jo paplitimo žemėlapį, atsižvelgiant į Rytų Europą ir Kaukazą (2.4 pav.). ).

Žemėlapyje rodomi didžiausi šio žymens dažniai tarp vakarų slavų (lenkų ir čekų), kur jis sudaro ketvirtadalį genofondo, o aplinkinėse populiacijose – šiek tiek sumažintas dažnis. Šio žymeklio paplitimas į vakarus apsiriboja gretimomis vokiečių populiacijomis (kurie tikriausiai asimiliavo vakarų slavų grupes, kurios anksčiau gyveno šiose srityse). Tačiau į rytus nuo haplogrupės aukšto dažnio zonos R-M458 plinta toli, užfiksuodamas visą rytų slavų arealą. Būdinga, kad ši rytinė ekspansija ryškesnė miško stepių zonoje (tarp ukrainiečių) nei miškų zonoje (baltarusų ir rusų). Dar įdomiau yra ribota plėtra į pietus: tarp pietinių slavų ir neslavų Balkanų tautų (vengrų, rumunų) šios haplogrupės dažnis yra mažas, o tai atitinka genetinę aukštų dažnių „ribą“. R-M458 ir geografinę Karpatų sieną. Į šiaurę nuo šios kalnų grandinės yra didžiausių haplogrupės dažnių zona, o į pietus nuo Karpatų dažnis smarkiai sumažėja. Matyt, Karpatai veikė kaip geografinė kliūtis genų srautui, neleidžianti šiai haplogrupei plisti į Balkanus. Atkreipkite dėmesį, kad, skirtingai nei R1 a apskritai bet kuriame Europos regione pasitaiko bent jau žemais dažniais, R- M458 paplitęs tik visoje Rytų Europoje, kaimyninėse Vakarų Europos ir Balkanų teritorijose, tačiau beveik visiškai nėra daugumoje Vakarų Europos – Iberijos pusiasalyje, Prancūzijoje, Britų salose, didelėse Apeninų ir Skandinavijos pusiasalių dalyse, taip pat nėra ir už jos ribų. Europa – o Vakarų Europoje – Azija ir Sibiras.
EUROPOS SKYRIUSR1 A-M558.
Antrasis Europos subvariantas R1 a nurodomas žymeklio pavadinimas R- M558 (2.5 pav.) Filogenetiškai ji yra broliška (dėl mtDNR tokiais atvejais įprasta sakyti „sesuo“, bet čia kalbama apie Y chromosomą) R- M458 . Abu šie žymekliai paskirstomi panašiai: R- M558 taip pat paplitęs daugiausia Rytų Europoje, taip pat mažiau paplitęs Balkanuose, žemais dažniais patenka į kaimyninius Vakarų Europos regionus ir kaip R- M458 , praktiškai nėra už Europos ribų. Šių potipių sritys taip pat sutampa didžiausio dažnio zonoje tarp Vakarų ir Rytų slavų. Pagrindinis skirtumas tarp paskirstytų modelių R- M458 Ir R- M558 susideda tik iš to, kad jie labiau apsiriboja vakarine ar rytine jų bendro arealo dalimi.

R- M458 turi tendenciją į vakarus nuo savo arealo. Tarp Vakarų slavų jis pasitaiko vidutiniškai 25% (t. y. ketvirtadalis genofondo), o tarp rytų slavų - 15%, nors vakarų baltarusiuose ir 2000 m. kai kurios Ukrainos populiacijos (bet ne Vakarų ir centrinės).
R- M558 linkęs į rytus nuo jų bendro arealo. Jis pasireiškia dažniau nei 30% abiejose pietų rusų populiacijose, tirtose dėl šio žymens; yra 20-25% tarp baltarusių, lenkų, slovakų, vakarų ukrainiečių; mažėja žemiau (arba žymiai žemiau) 20% tarp centrinių rusų, čekų, vidurio ir rytų ukrainiečių bei čekų. Tačiau R- M558 tirta žymiai mažiau populiacijų nei R- M458, ir todėl jo paplitimo žemėlapis ateityje dar gali būti gerokai patobulintas. Matome, kad net šis pagrindinis skirtumas tarp paskirstymo modelių R- M458 Ir R- M458 yra išreikštas mažais dažnių skirtumais ir turi gana sudėtingą geografiją.
Šis pavyzdys aiškiai parodo, kad filogenetinės skiriamosios gebos padidėjimas ne visada padidina geografinę skiriamąją gebą: nors ir Europos šakoje R1 a ir buvo galima nustatyti du potipius, tačiau jų geografinis pasiskirstymas labai panašus. Matyt, jie atsirado giminingose populiacijose ir išplito kartu tų pačių migracijų metu. Arba, net atsiradusios skirtingose populiacijų diapazono dalyse, kurios intensyviai keitėsi viena su kita, jos išplito visame diapazone. Žinoma, būtina padidinti filogenetinę skiriamąją gebą, ir, kaip taisyklė, nustatyti potipiai turi aiškesnę geografinę vietą nei visa šaka; kito lygio potipiai turi dar siauresnę pasiskirstymo zoną ankstesnio lygio potipių zonose ir pan. Tačiau šis geografinis aiškumas priklauso ne tik ir ne tiek nuo filogenetinės skiriamosios gebos lygio, bet ir nuo genofondo struktūros, nuo istoriškai nurodyto izoliacijos ir kryžminimosi santykio, tai yra, populiacijos genetikos požiūriu, nuo populiacijos pusiausvyros. genetinis dreifas ir migracija.
Jei ši pusiausvyra bus nukreipta į izoliaciją (kaip Kaukaze ar Sibire), tada haplogrupių dažnis tarp populiacijų labai skirsis, haplogrupės turės geografiškai siauras sritis, kurios mažai persidengs viena su kita, o haplogrupės pasieks labai aukštus dažnius savo „savo“ “ populiacijos . Jei balansas bus perkeltas migracijų link, tai dažniai taip smarkiai nesiskirs, sritys bus gana plačios ir persidengs viena su kita, o subhaplogrupės nepasieks per aukštų dažnių, nes likusiai genofondo daliai atstovaus kitos haplogrupės, kurių plotai sutampa su ta pačia teritorija. Visos šios savybės aiškiai matomos pavyzdyje R- M458 Ir R- M558 , nes Europai ypač būdingas balanso poslinkis į migraciją, o ne į izoliaciją.
EUROPOS SKYRIUSR1 A-Z284.
Tačiau yra dar vienas papildomas variantas R1 a– haplogrupė R- Z284 (2.6 pav.) turi visiškai kitokį raštą, kurį galima pavadinti „skandinavišku“ arba „šiaurės europiečiu“. Šiuo metu turimais duomenimis, Norvegijoje jo dažnis siekia 20%, Danijoje – 7%, Anglijoje ir Švedijoje – po 3% ir už Šiaurės Europos ribų. R- Z284 rasta tik keliuose pavyzdžiuose. Šie dažniai tikrai bus pakoreguoti, kai bus tiriamos naujos populiacijos ir didinamas jau tirtų tautų imčių dydis, tačiau bendras ryšys su Šiaurės Europos populiacijomis jau neabejotinas.

HAPLOGRUPĖR1 b(BENDRAI)
Haplogrupė R1 b, „broliškas“ haplogrupės atžvilgiu R1 a ir antras pagal dažnumą Europoje, taip pat turi platų diapazoną (2.8 pav.), tačiau pagrindinė jos didžiausių dažnių zona yra Vakarų Europoje. Svarbus indėlis į Europos genofondo tyrimą buvo naujų informatyvių SNP žymenų atradimas vienoje iš haplogrupės šakų. R1 b- haplogrupės R-M269(Žemėlapiuose pažymėta naudojant filogenetiškai artimą žymeklį kaip R-L265 ). Šį didelio masto tyrimą atliko didelė tarptautinė komanda, dalyvaujant autoriui, ir paskelbta m.
R1 b apskritai ( R- L10 , ryžiai. 2.8.) aiškiai parodo, kad tai yra pagrindinis komponentas, sudarantis daugiau nei pusę Vakarų Europos Y-chromosomų genofondo. Tik pietinės Apeninų pusiasalio dalies gyventojai turi sumažintus šios haplogrupės dažnius, nes tarp jų vyrauja Viduržemio jūros haplogrupės, o net Skandinavija pagal haplogrupių geografiją nepriklauso Vakarų Europai, o sudaro savarankišką šiaurės sritį. , kurioje vyrauja kita haplogrupė - aš1 .
Haplogrupė R1 b net labiau būdinga Vakarų Europai nei R1 a būdingas Rytams, nes R1 b daugelyje populiacijų (Britų salose, baskuose ir daugelyje kitų Iberijos pusiasalio populiacijų) jis sudaro net ne pusę, o daugiau nei du trečdalius genofondo. Apskritai haplogrupių tendencija R1 b- didžiausi dažniai baskuose ir laipsniškas dažnio mažėjimas į rytus nuo Iberijos pusiasalio - labai panašus į klasikinio žymeklio geografiją Rh-d(Rh faktorius). Dažnio padidėjimas Rytų Europoje rodo padidėjusį dažnį R1b kai kuriose baškirų populiacijose, žinomose iš [Lobov, 2009]. Tačiau mūsų komanda dabar baigia išsamų Y chromosomos įvairovės tyrimą baškirų populiacijose (daugiau nei 1000 mėginių). Šis tyrimas parodė, kad dažnis R1 b yra aukštas tik keliose klanų grupėse, tuo tarpu daugumai baškirų klanų grupių būdingos kitos haplogrupės ir dažniai R1 b jų maži. Todėl purslai R1 b Europos rytuose atspindi ne visos Europos modelį, o tik vietinius atskirų bendrinių Eurazijos stepių populiacijų grupių kilmės modelius.
Nors R1 b aukštus dažnius pasiekia daugiausia Vakarų Europoje, žemėlapyje (2.8 pav.) matyti, kad jis (kaip R1 a) yra paplitęs visur, užimantis visą žemėlapio diapazoną ir tęsiasi toli už savo sienų visoje Eurazijoje. Manoma, kad R1 b yra Vakarų Azijos kilmės, o įvairios jos atšakos išplito po įvairias Senojo pasaulio vietas, tačiau ypač „pasisekė“ tik atšakai, išplitusiai į Europą ir joje pasiekusiai tokius reikšmingus dažnius.

Paveiksle pavaizduotos tik šakos, kurios detaliai išnagrinėtos naudojant genogeografinius žemėlapius; diagramos struktūra atitinka pilną haplogrupės R1b-L10 medį pagal ISOGG versiją 2015 m. spalio mėn.

HAPLOGRUPĖS SUBVARIANTAIR1 b
Žemėlapiai pav. 2.9, 2.10 ir 2.11 rodo tris pagrindines šakas R1 b(2.7 pav.), kurias – su daugybe išlygų – galima vadinti Afrikos, stepių Eurazijos ir Vakarų Europos haplogrupėmis. Iš tiesų, kai žiūrime į žemėlapį R1 b apskritai (2.8 pav.) matome jos buvimą ir Šiaurės Afrikoje.
AFRIKOS FILIALASR- V88.
Žemėlapis R- V88 (2.9 pav.) tik rodo dažnius, nors ir žemus (1-6%, tik vienoje populiacijoje padidėjo iki 26%), bet visoje Šiaurės Afrikoje (nuo Maroko iki Egipto), ir beveik visišką šios haplogrupės nebuvimą. Europoje ir Azijoje. Tai yra „afrikietiška“ dalis R1 b– tai beveik išimtinai filialas R- V88 . Jis pasiekia maksimumą toli už žemėlapio ploto – Centrinėje Afrikoje.

STEPĖS ŠAKALASR- M478 .
Kitos šakos žemėlapis - R- M478 – rodo jo ryšį su Eurazijos stepių populiacijomis (2.10 pav.). Jo didžiausių dažnių zona taip pat didžiąja dalimi yra už žemėlapio zonos ribų – erdvėse nuo Uralo iki Altajaus. Tačiau, kaip matote žemėlapyje, atskiromis „kalbomis“ ši haplogrupė pasiekia Kaukazą, apsiribodama Pietų Uralo ir Ciskaukazijos stepių regionais ir sudaro tik 1-10%, retai iki 20 populiacijų. % genofondo.


VAKARŲ EUROPOS SKYRIUSR- L265.
Trečioji iš pagrindinių haplogrupės šakų R1 b- haplogrupė R-M269, arba, kaip čia pažymime žemėlapiuose, R- L265 — sudaro pagrindinę (dažniu ir plotu) R1b dalį. Todėl jo paplitimo žemėlapis (2.11 pav.) apskritai atkartoja žemėlapį L10 , tik be afrikietiškos jo arealo dalies. Ši haplogrupė yra suskirstyta į daugybę subhaplogrupių, iš kurių daugelis yra būdingos tam tikrai šakos diapazono sričiai. Sutelkime dėmesį į dvi pagrindines šakas viduje R- L265 : Vidurio Europos haplogrupė R- M405 ir jos Iberijos-Britanijos „brolis“ R- P312 su visomis savo „filialinėmis“ šakomis (2.12. - 2.17. pav.).

Centrinės Europos filialas R-M405 BENDRAI.
Haplogrupė R-M405(2.12 pav.) aptinkama beveik visoje Vakarų, Vidurio ir Šiaurės Europoje (išskyrus pietinius Iberijos ir Apeninų pusiasalio regionus, taip pat Balkanų pusiasalį pietuose, Suomiją ir Kolos pusiasalį šiaurėje). ), taip pat vakarinėje Rytų Europos dalyje . Tačiau esant maksimaliems dažniams (18-36%), haplogrupė R-M405 randama daugiausia Vidurio Europos (Vokietijoje, Šveicarijoje, Belgijoje, Olandijoje, Danijoje ir pietų Britanijoje) gyventojų. Vidinių haplogrupės subvariantų atsiradimo ir identifikavimo data R-M405 pagal www.yfull.com yra maždaug prieš 5 tūkstančius metų (prieš 4,4-5,3 tūkst. metų), t.y. datuojamas bronzos amžiumi. Atrodo, kad jau tuo metu vyko migracijos procesai, lėmę haplogrupės plitimą R- M405 į įvairias Europos dalis. Gali būti, kad duomenys apie haplogrupę R- M405 nurodo vieną iš indoeuropiečių kalbų kalbėtojų plitimo Europoje epizodų. Į vieną iš haplogrupių nešėjų pasiskirstymo krypčių R- M405, galbūt tai rodo vienos iš jos pošakių – haplogrupės – geografija R- M467.

„VOKIEČIŲ-BRITŲ“ SKYRIUSR- M467.
Kuklus haplogrupės pasiskirstymas R- M467 (2.13 pav.) - tiek savo dažniais (nuo daugiausiai 4% rytų Vokietijoje iki maždaug 3% šiaurės ir pietų Vokietijoje, Olandijoje ir pietų Britanijoje iki 0,5% Prancūzijoje), tiek pagal geografiją (ne didžiausios, pagal Europos standartus, teritorijos aplink Šiaurės jūrą) - vis dėlto tai domina. Haplogrupės geografija R- M467 siūlo jos garsiakalbių migraciją iš šiuolaikinės Vokietijos teritorijos palei Šiaurės jūrą į Britų salų pakrantes. Pažintys haplogrupei R- M467, www.yfull.com pateiktame tinklalapyje nurodoma, kad jis atsirado maždaug prieš 4,7 tūkst. metų (riboje prieš 4,1-5,4 tūkst. metų), o išsiplėtimo (skirstymo į pošakius) laikas po tūkstančio metų yra maždaug prieš 3,4 tūkst. (riboje nuo 2,8-4,0 tūkst. metų). Palyginus haplogrupės diapazoną R- M467 žemėlapyje su nurodytomis atsiradimo ir paplitimo datomis galime daryti prielaidą, kad tokios situacijos atsiradimo modeliai yra du. Pirmasis – kai migracijos procesai iš centrinių šiuolaikinės Vokietijos regionų teritorijos bronzos amžiuje perėjo Šiaurės jūros pakrante į Britų salas ir paliko pėdsaką šiuolaikiniame genofonde. Antrasis – kai ši haplogrupė galėjo būti pasiskirstyta kiek plačiau į pietus, vakarus ir rytus, tačiau dėl to, kad ten atvyko kita populiacija, haplogrupės dažnis ir taip žemas. R- M467 pietuose sumažėjo iki verčių, kurių beveik neįmanoma nustatyti esant dabartiniams imties dydžiams.
PIRENĖJŲ IR DRIČIŲJŲ ŠALIŲ SKYRIUSR- P312 IŠ VISO.
Haplogrupių pasiskirstymas R- P312 (2.14 pav.) yra įdomus tuo, kad iš esmės atspindi bendruosius pasiskirstymo tarp Europos, kaip jos protėvių haplogrupės, ypatumus. R- L265 , ir visa haplogrupė R1 b apskritai. Aukščiausi haplogrupių dažniai R- P312 pačiuose Europos vakaruose (Iberijos pusiasalyje, Vakarų Prancūzijos teritorijoje, Britų salose), palaipsniui mažėjant dažniui į rytus ir vietiniu antplūdžiu pietų Urale - tai beveik tos pačios savybės, kurios buvo aprašytos aukščiau, kai apibūdinantis haplogrupės plotą R1 b apskritai. Tačiau laikytis bendros tendencijos nereiškia visiško panašumo. Taigi didžiojoje Vakarų Europos dalyje yra didžiulė raudonai violetinių atspalvių sritis (dažniai 50–75%), kuri anksčiau buvo pastebėta haplogrupių žemėlapiuose. R1 b- L10 Ir R- L265 (2.8 pav. ir 2.11 pav.), haplogrupių žemėlapyje R- P312 (2.14 pav.) virsta vietiniu dažnio pliūpsniu (iki 75%) Britų salų šiaurėje ir geltonai raudonų atspalvių sritimi (dažnis 30-50%) Pirėnų pusiasalyje ir Prancūzijoje. Pietų Uralo raudonai violetinis bendro dažnio pliūpsnis R1 b haplogrupių žemėlapyje R- P312 yra sumažintas iki mažos „akies“ šiaurės baškirų taške, kurį iš visų pusių supa žemo dažnio regionai (ne daugiau kaip 3%).
Haplogrupė R-P312, taigi apima didelę dalį visos haplogrupės įvairovės R- L265 , taigi jo diapazonas. Galima daryti prielaidą, kad šiuo atveju haplogrupėje R- P312 Taip pat yra keletas vietinių padalinių su siauresnėmis sritimis. Ir ši prielaida pasiteisina: šiandien žinomos penkios didelės šakos, kurios savo ruožtu skirstomos į keliasdešimt gilesnių. Tačiau mes sutelksime dėmesį tik į tris šakas R- P312 , kurio paplitimas Europoje gerai ištirtas: haplogrupė R-M167 ir jo subvariantas R- M153 , taip pat haplogrupė R- M222 (2.15-2.17 pav.).

„BASCO-ROMAN“ HAPLOGRUPĖR- M167.
Tai yra haplogrupės pavadinimas R- M167 sąlyginai: jos geografija (2.15 pav.) apima Iberijos pusiasalio ir Vakarų Prancūzijos gyventojus, nedidelę teritoriją Vokietijos pietuose ir vieną „akelį“ Olandijoje, taip pat teritoriją Vakarų Juodosios jūros regione (rumunai). ir bulgarai). Taigi didžiojoje haplogrupės arealo dalyje gyvena tautos, kalbančios romanų (ispanų, portugalų, prancūzų ir rumunų) indoeuropiečių kalbų šeimos ir baskų kalbų šeimos kalbomis. Tačiau pietų vokiečiai ir bulgarai, kurie taip pat turi haplogrupę R- M167 , kalba kitų indoeuropiečių šeimos grupių kalbomis.
Didžiausių haplogrupių dažnių sritis R- M167 (gelsvai žalsvi tonai 2.15 pav., haplogrupių dažnis nuo 6% iki 25%) apima Pirėnų pusiasalio ir pietvakarių Prancūzijos teritoriją. Ta pati sritis yra vienintelė dukterinės haplogrupės paplitimo teritorija R- M167 – subvariantas R- M153 (2.16 pav.).

„BASKŲ“ ŠAKALASR- M153.
Kaip matyti žemėlapyje (2.16 pav.), haplogrupė R- M153 pasiekia maksimalų dažnį (beveik 16%) tarp baskų, yra perpus mažiau paplitęs Ispanijos Pirėnų populiacijoje, o likusį diapazoną apima labai žemais dažniais (1-3%). Dėl šios geografijos haplogrupė R- M153 galima sąlygiškai vadinti „basku“ ir daryti prielaidą, kad ji atsirado ir išplito daugiausia šios tautos populiacijose. Šios haplogrupės kilmės amžius pagal www.yfull.com yra apie 2,5 tūkstančio metų (svyruoja nuo 3500 iki 1500 metų). Kitaip tariant, ši haplogrupė greičiausiai atsirado Pirėnų kalnų populiacijose mūsų eros aušroje ir sugebėjo išplisti abiejose kalnų grandinės pusėse (ir efektyviau į pietus nuo Pirėnų) dėl vietinių migracijų. .


D. Britanijos filialasR- M222.
Haplogrupę galima laikyti stebinančia, tik ne jos diapazono platumu, o būtent kompaktiškumu. R- M222 (2.17 pav.). Šios haplogrupės paplitimas beveik vien tik Britų salose rodo vietinę kilmę. Remiantis www.yfull.com skaičiavimais, haplogrupės išplėtimas R- M222 datuojamas tik 1,8 tūkst. metų (prieš 1,3-2,4 tūkst. metų). Kitaip tariant, haplogrupė Britų salų populiacijose atsirado II tūkstantmetyje prieš Kristų, o ją paskleidusios migracijos įvyko mūsų eros pradžioje, tai yra maždaug romėnų laikais.
Haplogrupių filogeografijos pavyzdys R-L265 parodo, kaip išsami genetinė informacija yra paslėpta kiekvienoje Y chromosomos haplogrupėje. Galime tik tikėtis, kad kaupiantis duomenims apie pilnus genomus, kitose haplogrupėse bus aptikti informaciniai žymenys.
INDIJOS HAPLOGRUPĖR2 (R- L266).
Haplogrupė R2 (R- L266 ) kilęs iš tos pačios šaknies kaip ir haplogrupė R1 , dėl kurios susidarė haplogrupės R1 a Ir R1 b. Haplogrupių pasiskirstymo žemėlapis R2 (R- L266 ) (2.18 pav.) rodo, kad visos Europos gyventojams R2 nėra tipiškas: itin žemu dažniu (mažiau nei 3%) pasitaiko tik Apeninų pusiasalio šiaurėje ir Sardinijoje, tarp turkų gyventojų prie Dardanelų sąsiaurio, vidurio Anatolijoje ir Užkaukazėje. Su šiek tiek didesniu 5% haplogrupės dažniu R2 (R- L266 ) pastebėta pietryčių Turkijoje, o tarp kalmukų – 6 %. Tokia menka haplogrupės geografija kartografuotoje srityje turbūt nestebina: juk pagrindinė sritis R2 (R- L266 ) apima tolimas Indijos pusiasalio ir Centrinės Azijos teritorijas, o žemais dažniais haplogrupė pasiekia Kiniją rytuose ir Pietvakarių Aziją jos arealo vakaruose.

Tęsdamas temą apie didelį informacijos turinį nustatant potipius ir analizuojant jų geografinį pasiskirstymą, leiskite jums priminti klasikinį darbą, kuriame haplogrupė aš-M170 buvo filogenetiškai suskirstytas į tris potipius. Paaiškėjo, kad šie potipiai turi visiškai skirtingas geografines vietas : haplogrupė I1-M253(žemėlapiuose — I-L118) apsiriboja Šiaurės Europa, I2a-P37– į Balkanų regioną, ir I2b-M223(žemėlapiuose - I-L35) daugiausia lokalizuotas Šiaurės Vakarų Europoje. Būtent po šio ir daugybės panašių darbų paaiškėjo, kad naujų SNP žymenų, dalijančių haplogrupę į pošakius, atradimas yra pagrindinis būdas padidinti Y chromosomų analizės informacijos turinį. Tačiau ilgą laiką pažanga šiuo keliu buvo lėta ir tik įvedus visišką Y chromosomos sekos nustatymą, nauji žymenys ėmė kauptis kaip lavina. Taigi ilgą laiką haplogrupėje nebuvo įmanoma atrasti naujų SNP žymenų aš, arba naujai atrasti SNP žymenys nenustatė naujų šakų su aiškia geografija. Pavyzdžiui, darbe buvo atrasti nauji žymekliai ir pertvarkyta haplogrupių šakų topologija I-M170, tačiau buvo išsaugotos tos pačios trys pagrindinės šakos, nors ir naujais pavadinimais, o naujai atrasti variantai buvo reti ir nepasižymėjo aiškiomis geografinėmis tendencijomis.
„SKANDINAVIJA“ HAPLOGRUPĖ I-L118.
„Skandinaviškos“ haplogrupės paplitimo žemėlapis I-L118(2.19 pav.) rodo maksimalūs jo dažniai ne tik geografinėje Skandinavijoje, bet ir su ja istoriškai glaudžiai susijusiose teritorijose: Danijoje, Škotijoje, vakariniuose Suomijos regionuose. Todėl į šiaurę nuo Volgos esantis Rusijos gyventojų vidutinio dažnio regionas, kuris daugumos slavų populiacijų žemų dažnių fone išsiskiria, yra įdomus. Negalima atmesti, kad tai atspindi istorinį kontaktą su normanų populiacijomis. Nors šie ryšiai buvo intensyvesni šiaurės vakarų rusų populiacijoms (kelyje „nuo varangų iki graikų“), šiaurės vakarų teritorijos buvo gana tankiai apgyvendintos, todėl „varangiečių“ įtaka galėjo būti reikšmingesnė mažesniems. Trans-Volgos regiono gyventojų.

"BALKAN" HAPLOGRUPĖ I-P37
„Balkanų“ haplogrupės paplitimo žemėlapis I-P37(2.20 pav.) rodo aukštus jo dažnius ir Italijoje. Apskritai ši haplogrupė yra paplitusi beveik visoje Europoje, išskyrus Skandinaviją. Rytų Europoje sklandus mažėjimo dažnio gradientas iš pietvakarių (Ukraina) į šiaurę ir rytus labai primena pirmojo pagrindinio komponento žemėlapį, pagrįstą klasikiniais genetiniais žymenimis [Rychkov ir kt., 2002]. Taip pat verta atkreipti dėmesį į gana ryškų šios haplogrupės ploto sutapimą su geografinėmis Europos sienomis - dažnai Juodosios jūros regione haplogrupė yra labai reta Šiaurės Kaukaze ir beveik nėra Užkaukazėje, o su maksimaliais dažniais Balkanuose jis labai retas kaimyninėje Mažojoje Azijoje.

„VARIAGO-GRAIKIJOS“ HAPLOGRUPĖ I-L35
Šiaurės ir Baltijos jūrų pakrantėse ir šiek tiek į vidų nuo jų – taip būtų galima apibūdinti haplogrupės geografiją aš-L35, jei ne dar du papildymai šiaurės rytuose ir pietryčiuose. Nors maksimalus haplogrupės dažnis aš-L35 ( 17 proc. pažymėta Švedijoje (2.21 pav.), retesnio haplogrupės pasireiškimo zona (3-7 proc.) apima šiaurinę Vidurio Europos dalį, pietryčiuose atskira labai žemo dažnio zona (1- 3%) apima Balkanų pusiasalį ir vakarinę Mažosios Azijos dalį, o šiaurės rytuose pasireiškia Tverės ir Kostromos regionų rusų populiacijomis. Galima pastebėti, kad haplogrupės diapazonas aš-L35 — haplogrupė yra gana vidutinė ir vienalytė dažnio reikšmėmis (daugumoje plotų dažnis svyruoja nuo 2-3% iki 7%, tik Švedijoje pikas yra iki 17%) - ji jungia kelis kraštutinius Europos regionus: Skandinavija, šiauriniai Rytų Europos regionai ir Pietų Europos rytai. Visai įmanoma, kad haplogrupės geografija aš-L35 iš tikrųjų atspindi vieną iš genetinių prekybos kelių tarp Europos šiaurės ir pietų pėdsakų. Ne mažiau tikėtina, kad kiekviena iš šių zonų yra susijusi su savo šios haplogrupės subvariantu.

Haplogrupė N Europos regione jį atstovauja dvi didelės šakos – haplogrupės N1c Ir N1b(toliau ir žemėlapiuose N-M46 Ir N-P43 atitinkamai). Nors bendrosios šių dviejų haplogrupių sritys Europoje persidengia, jų didžiausių dažnių zonas riboja šiaurėje Pečoros upė, o pietuose – Uralo kalnagūbris: iš vakarų nuo šios sąlyginės „sienos“ lieka zonos padidėjęs haplogrupės atsiradimas N-M46, o iš rytų – maksimalių dažnių zona N-P43. Panagrinėkime šių dviejų „broliškų“ haplogrupės atšakų geografiją Nšiek tiek detaliau.
ŠIAURĖS EUROPOS VAKARŲ HAPLOGRUPĖ N-M46
Haplogrupių pasiskirstymo žemėlapis N-M46(2.22 pav.) rodo didžiausi jo dažniai tarp suomių. Jo dažnis yra aukštas ir kitose šiaurės rytų Europos populiacijose. Nors apskritai atrodo, kad ši haplogrupė apsiriboja finougrų populiacijomis, ji taip pat paplitusi tarp šiaurės rusų (kas, matyt, paaiškinama ikislavų gyventojų asimiliacija, kaip bus aprašyta tolesniuose skyriuose). Įdomiausia sritis atrodo didelis šios haplogrupės dažnis tarp baltų (latvių ir lietuvių). Šiuo atveju genetinės ribos gerai sutampa su kalbinėmis: baltų ir vakarų slavų plotų riba sutampa su ryškiu haplogrupės dažnio „uoliu“. Panašus vaizdas stebimas pasienyje su baltarusiais.

„ŠIAURĖS URAL“ HAPLOGRUPĖ N-P43
Haplogrupių pasiskirstymo žemėlapis N-P43(2.23 pav.) taip pat atskleidžiamas šiaurinis, bet gerokai siauresnis arealas. Didžiausi šios haplogrupės dažniai stebimi kraštutiniuose Europos šiaurės rytuose ir sklandžiai pereina prie pasaulinio šios haplogrupės maksimumo Vakarų Sibire. Esant vidutiniams dažniams, ši haplogrupė yra paskirstyta visame Urale ir Urale. Pastebimas sklandus mažėjančio dažnio gradientas į vakarus ir pietus, todėl Centrinės Rusijos ir Volgos žemupio populiacijose ši haplogrupė vis dar randama (nors ir minimaliais dažniais), o kitose Europos populiacijose jos beveik visiškai nėra.

Dauguma aukščiau aprašytų haplogrupių daugiausia apibūdino sąlyginai šiaurinę Europos žemyno pusę. Atsižvelgiant į šios haplogrupės geografiją - E– dėmesį nukreipsime į pietus, į Viduržemio jūros pakrantes. Keturios pagrindinės haplogrupės atšakos yra Europoje E: E- L142 Ir E- V22 (grįžta prie bendros šaknies - haplogrupės E-M78), E- M81 Ir E- M123 .
RYTŲ VIDURŽEMIO JŪROS HAPLOGRUPĖE- L142
Haplogroup žemėlapis E- L142 (2.24 pav.) šiek tiek primena haplogrupių žemėlapį I-P37(2.20 pav.) – abiejų šių haplogrupių didžiausi dažniai apsiriboja Balkanų pusiasalyje. Jų modeliai taip pat labai panašūs: dažnis mažėja į šiaurę, vakarus ir rytus. Pagrindiniai šių dviejų haplogrupių tendencijų skirtumai yra, pirma, mažesnis dažnis E- L142 (daugelyje teritorijų jis yra „prastesnis“ I-P37 per žemėlapio mastelio intervalą) ir, antra, pasiskirstyme E- L142 ne tik Europoje, bet ir visoje Viduržemio jūroje, įskaitant Artimuosius Rytus ir šiaurines Afrikos dalis. Balkanų viduje I-P37 ypač paplitusi tarp bosnių ir kroatų, ir E- L142 vyrauja pietuose – tarp serbų, albanų ir graikų.

„EGIPTIETĖ“ HAPLOGRUPĖE- V22
Dvi mažos „akys“ Iberijos pusiasalio ir Apeninų pusiasalio šiaurėje ir pietuose - teritorijos, kuriose yra haplogrupės dažnis E- V22 iki 5%, nedidelė teritorija Mažosios Azijos šiaurės vakaruose (prie Stambulo), kurios dažnis iki 6%: tai galėtų užbaigti haplogrupės geografijos aprašymą. E-V22 geografinėse Europos ribose (2.25 pav.). Tačiau haplogrupės geografija E-V22 Europoje daugiausia eina Viduržemio jūros pakrante, todėl verta atsekti jo paplitimą toliau į pietus. Žemėlapis pav. 2.25 rodo sklandų haplogrupės dažnio padidėjimą E- V22 dviejuose priešinguose regionuose: pietvakariuose ir pietryčiuose nuo Viduržemio jūros. Viduržemio jūros pietvakariuose prie Gibraltaro sąsiaurio krantų ir giliai į Maroką haplogrupės dažnis E-V22 pakyla iki 7 proc. Pietryčių Viduržemio jūros dalyje haplogrupės dažnio padidėjimas E- V22 Jis pastebimas nuo Mažosios Azijos į pietus - per Levantą iki Nilo deltos, kur siekia 14%, ir dar toliau į pietus palei Nilą, kur tarp Baharijos oazės egiptiečių arabų jis pasiekia didžiausią pasaulyje 22%. . Iš šių dviejų neeuropinių haplogrupės paplitimo regionų E- V22, Tikėtina, kad būtent pastaroji (su didesniu dažniu ir platesniu pasiskirstymu) buvo pagrindinis haplogrupės pasiskirstymo šaltinis. E- V22 į Europą.

SAHAROS HAPLOGRUPĖE- M81
Jei dvi ankstesnės haplogrupės E šakos yra E- L142 Ir E-V22 - daugiausia pademonstruotas pasiskirstymas šiaurinėje ir rytinėje Viduržemio jūros pakrantėje, kartais gilėjant į šiaurę ir rytus nuo žemyno, tada trečiosios šakos geografija - haplogrupės E- M81- daugiau seka pietinę Viduržemio jūros dalį. Haplogrupių pasiskirstymo žemėlapis E- M81 pav. 2,26, nors tai rodo pastebimą haplogrupės buvimą Artimuosiuose Rytuose (dažnis iki 20%), tačiau pagrindinė aukštų dažnių zona yra šiaurės vakarų Afrikoje. Nuo šio maksimumo buvo pastebėtas sklandus dažnio sumažėjimas šiaurės kryptimi per Gibraltaro sąsiaurį su vidutiniu haplogrupės pasiskirstymu (2-10%). E- M81 Iberijos pusiasalio ir pietvakarių Prancūzijos teritorijoje, siauros žemo dažnio zonos (ne daugiau kaip 3%) Apeninų ir Balkanų pusiasalyje, taip pat Mažosios Azijos šiaurės rytuose.


PRIE AZIJOS HAPLOGRUPĖSE- M123
Haplogrupė E- M123 pasiekia maksimumą Vakarų Azijoje (20-25 proc.), arčiau Europos (Mažojoje Azijoje) sumažėja iki 15 proc. Pačioje Europoje žemais dažniais išplitusi šiauriniuose Viduržemio jūros pakrantėse, pavieniais lopais pasirodo Vidurio Europoje (2.27 pav.).
Dviejų haplogrupių atšakų haplogrupių pasiskirstymo žemėlapiai G- P15 – haplogrupes G- P303 irG- P16 – apskritai jų pasiskirstymas panašus ir net visos Europos žemėlapio masteliu aišku, kad abu daugiausia apsiriboja Kaukazu. Tačiau jie pasiekia didžiausią dažnį skirtingose Kaukazo regiono dalyse: haplogrupėje G- P303 vyrauja kraštutiniuose šiaurės vakaruose tarp Juodosios jūros šapsugų ir haplogrupės G- P16 - Centriniame Kaukaze tarp osetinų-ironiečių (2.28 pav. ir 2.29 pav.). Tai visiškai patvirtina atskiras Kaukazo genofondo tyrimas.

VAKARŲ KAUKAZIO HAPLOGRUPĖG- P303
Žemėlapyje parodyta, kad haplogrupė G- P303 (2.28 pav.) paplitęs ir Artimuosiuose Rytuose, o Europoje aptinkamas daugiausia pietrytiniuose jos regionuose. Aukšto dažnio haplogrupių zonos G- P303 driekiasi palei rytinę Juodosios jūros pakrantę ir yra tik Vakarų Kaukazo tautų, daugiausia abchazų-adigėjų kalbinės grupės, populiacijos. Su žemesniu dažniu (iki 10%) haplogrupė G- P303 paplitęs Šiaurės Juodosios jūros regione (Krymo pusiasalio teritorijoje ir Azovo srityje), žemo dažnio regionas (3-5%) tęsiasi toliau į šiaurę į Ukrainos miškų stepių zoną. Vietinis haplogrupių dažnio padidėjimas G- P303 iki 13% taip pat stebima Volgos regione tarp Mordovijos moksų gyventojų ir yra apsupta žemų dažnių regiono.

Centrinės kaukazo HALOGRUPĖG- P16
Haplogrupės geografija G- P16 (2.29 pav.) siauresnis lyginant su savo „broliu“ šaka G- P303 . Ji apima tik Kaukazo regioną ir Mažąją Aziją. Šioje srityje haplogrupės dažnis G- P16 staigiai nukrenta nuo didžiausių osetinų (73% tarp osetinų-ironiečių ir 56% tarp osetinų-digoriečių) iki 10-20% šiaurės vakarų Kaukazo tautų (abchazų, čerkesų, balkariečiai ir karačajai), o po to sumažėja iki minimumo tarp turkų, armėnų ir azerbaidžaniečių.
Haplogrupė J atstovaujama dviejų pagrindinių šakų: J- L255 (J1) Ir J-L228 (J2), iš kurių antrasis yra labiau paplitęs Europoje. Nors abi šios šakos yra kilusios iš Artimųjų Rytų ir atspindi senovinius ryšius tarp Europos gyventojų ir daug pietesnių Eurazijos regionų gyventojų, prasminga atskirai apsvarstyti kiekvienos iš šių linijų geografiją kaip visumą ir kiekvienos iš jų. Europoje labiau paplitusios haplogrupės vidinės atšakos J-L228 (J2).

HAPLOGRUPĖJ- L255 (J1): nuo Vakarų Azijos iki Rytų Kaukazo viršūnių
Haplogrupių pasiskirstymas J- L255 (2.30 pav.) kartografuotoje teritorijoje daugiausia apima jos pietrytinius regionus: Kaukazą, Mažąją Aziją, Šiaurės Juodosios jūros regioną, Balkanų pusiasalį; mažos zonos – Apeninų pusiasalio ir Prancūzijos pietuose, Pirėnų pusiasalio pietvakariuose. Didžiausių dažnių regione - Kaukaze - haplogrupė J- L255 pabrėžia Rytų Kaukazo tautas, kur ji sudaro liūto dalį viso genofondo. Didžiausia haplogrupė J- L255 pasiekia Dagestaną (tarp kubachų, darginų, tabasaranų ir avarų jo dažnis svyruoja nuo 63 iki 98 proc.), o tarp lezginų – iki 44 proc., tarp čečėnų – iki 21 proc. Mažosios Azijos teritorijoje haplogrupės dalis J- L255 svyruoja nuo 5% iki 15%.
HAPLOGRUPĖJ-L228 (J2) BENDRAI
Haplogrupių pasiskirstymo žemėlapis J-L228 (J2) piešia kitokį paveikslą (2.31 pav.). Skirtingai nuo daugelio aukščiau aptartų haplogrupių, J-L228 (J2) platinami aukštais dažniais ne Europoje, o Artimuosiuose Rytuose ir Šiaurės Afrikoje. Žinoma, jo arealas taip pat apima pietinius Europos regionus, besiribojančius su šiais regionais: Ispaniją (ypač jos pietinius regionus), Italiją (taip pat ypač pietus) ir Balkanų pusiasalio pietus. Tačiau ši haplogrupė pasiekia didžiausią dažnį (žemėlapyje raudona) kai kuriose Šiaurės Kaukazo populiacijose. Kaip buvo parodyta mūsų tyrime ir darbe, Kaukazo maksimumas apsiriboja Nakh grupės populiacijomis (čečėnais ir ingušais).


HAPLOGRUPĖJ- L152 IR JO SUBVARIANTASJ- M67
Haplogrupės geografija J- L152 Europoje yra gana plati: žemo dažnio zona driekiasi nuo Balkanų pusiasalio šiaurės vakarų per Vidurio Europą ir iki Iberijos pusiasalio. Aukštų dažnių zona (nuo 15%) apima Mažąją Aziją ir Kaukazą. Beveik pakartoja šį modelį, bet šiek tiek siauresnėje geografinėje sferoje ir paprastai mažesniais dažniais, jos „filialinės“ šakos - haplogrupės - pasiskirstymas. J-M67(2.32 pav.). Tačiau haplogrupė J-M67 savo dažniu labiau apsiriboja Kaukaze, sudarantis ne daugiau kaip 13% Mažosios Azijos teritorijos genofondo.
HAPLOGRUPĖJ- L282
Haplogrupių pasiskirstymas J- L282 Europoje (2.33 pav.) pastebimai skiriasi nuo ankstesnės šakos. Didžiulės žemų dažnių zonos fone išsiskiria trys sritys, kuriose yra haplogrupės dalis J- L282 genofonde padidinama iki 10-15 %: Fennoskandijos šiaurėje (tačiau šis maksimumas pagrįstas tik vienos populiacijos duomenimis), Volgos regione (Moksha populiacija mordoviečių) ir Balkanų pusiasalio šiaurėje (keli albanai). populiacijos).

RETOS HAPLOGRUPĖSL, K, T
HAPLOGRUPĖ L-M11
Haplogrupė L-M11(2.34 pav.) Europoje apsiriboja pietryčių regionais (išskyrus pavienį atvejį Belgijoje): Krymo pusiasalį, europinę Turkijos dalį ir Šiaurės Kaukazą. Be to, haplogrupės dažnis L-M11 palaipsniui didėja pietryčių kryptimi, pasiekdamas 14% Rytų Kaukaze tarp čečėnų ir 13% Mažosios Azijos šiaurės rytuose tarp turkų. Be to, jo dažnis didėja Artimuosiuose ir Viduriniuose Rytuose, pasiekdamas Indijos maksimumą jau toli už žemėlapio teritorijos.

HAPLOGROUP Q-M242
Haplogrupė Q-M242 Europoje jis praktiškai neaptinkamas, išskyrus keletą taškų su žemu dažniu (iki 3%) Vidurio ir Rytų Europoje, Krymo pusiasalyje ir rytinėje Mažosios Azijos pusėje (2.35 pav.). Lygus koridorius nuo Kaspijos jūros šiaurės vakarinės pakrantės toliau į rytus, palaipsniui didėjant dažniui, yra haplogrupės sritis Q-M242 pereina į Vidurinę Aziją ir Sibirą, kur pasiekia maksimumą.


HAPLOGRUPĖ T
Haplogrupių dažnis T-L206 Europos gyventojų neviršija 5% (2.36 pav.). Jo arealas Europoje yra kratinys ir vakaruose sumažintas iki trijų siaurų Pirėnų pusiasalio ir Sardinijos regionų, o rytuose iki zonos šiauriniame Juodosios jūros regione su perėjimu iš šiaurės rytų į Ukrainos miškų stepę, izoliuotą. atvejų Volgos regione tarp Kazanės totorių ir Šiaurės Kaukaze tarp Kubos nogajų. Už Europos ribų haplogrupės T dažnis yra vienodai mažas, tačiau jos arealas nebėra nevientisas, o beveik ištisinis, apimantis visą Mažąją Aziją, Artimuosius Rytus ir šiaurės rytų Afriką.
Taigi, Y-chromosomų kintamumo Europos populiacijose tyrimas, pagrįstas gausių nuosavų duomenų ir literatūros deriniu, patvirtino ir išaiškino išvadą, kad pagrindinis genofondo struktūrizavimo bruožas yra aiškus geografinių zonų nustatymas kurioje vyrauja sava haplogrupė. Tai įtikinamai rodo haplogrupių pasiskirstymo žemėlapiai (2.2.-2.36 pav.).
Tačiau pateikiant kiekvienos haplogrupės duomenis į atskirą žemėlapį, sunku atsekti haplogrupių sričių sutapimą arba, atvirkščiai, nepersidengimą. Todėl visų haplogrupių paplitimo zonas sujungėme viename žemėlapyje (2.37 pav.). Šiame paveiksle kiekvienos iš devynių pagrindinių Europos haplogrupių kontūrai griežtai atitinka teritoriją, kurioje šios haplogrupės dažnis yra didesnis nei 35%, t.y. daugiau nei trečdalis genofondo. Galima pastebėti, kad kiekviena haplogrupė iš tikrųjų užima savo diapazoną. Sritys, kurios lieka baltos, yra mažos – tai yra tos sritys, kuriose haplogrupių įvairovė yra didelė ir nė viena haplogrupė nepasiekia 35% lygio.
Gautas žemėlapis aiškiai parodo „galvosūkio principą“ – didelį Y chromosomos haplogrupių geografinį specifiškumą. Dėl šios priežasties kiekvienoje teritorijoje, nors ir yra daug haplogrupių, dominuoja tik viena ar dvi. Taigi Europos genofondas pasirodo tarsi iš galvosūkio sudarytas iš skirtingų haplogrupių vyraujančio pasiskirstymo sričių.

Kiekvienoje geografinėje Europos dalyje dominuoja viena haplogrupė, retai randama kitose dalyse. Žemėlapyje parodyta, kuriose Europos dalyse yra apsiribojusi kiekviena iš pagrindinių haplogrupių. Žemėlapis pagrįstas tiksliomis dažnio reikšmėmis. Zonos, kurių haplogrupių dažnis viršija slenkstinę reikšmę 0,35, rodomos spalvotai (tai yra, spalva paryškintose srityse šiai haplogrupei priklauso daugiau nei trečdalis genofondo).
Skliausteliuose pažymėkime, kad objektyvus šios haplogrupės susiejimas su slavų populiacijomis šioje labai ribotoje teritorijoje, deja, tapo viena iš priežasčių, kodėl „slaviška“ etiketė buvo priklijuota visai R1a haplogrupei, taip plačiai paplitusiai mėgėjų forumuose internete. Tačiau toks haplogrupės susiejimas – per visą jos Eurazijos arealą erdvėje ir išplėstą egzistenciją, neprilygstamą slavams laike – su slavų kalbine grupe, deja, ne tik ribojasi su pseudomokslu, bet ir kerta šią ribą.
Jei tarp šiuolaikinių Europos tautų Europoje galima išskirti keletą pagrindinių haplogrupių (daugiau nei 5%), tai, žinoma, yra R1b, R1a, I1, N1, I2, G2 šiaurėje ir J, E1b1, T pietuose. Europos, tada senovės Europos gyventojų padėtis yra visiškai kitokia.
Subkladas I2-M26 yra viena iš pagrindinių Y-chromosomų linijų Sardinijoje (siekia 40%) ir tarp baskų. Jis taip pat retai aptinkamas visoje pietvakarių Europoje: Ispanijoje, Prancūzijoje, Portugalijoje, Italijoje.
Subkladas I2-M423 būdingas Balkanų ir Karpatų populiacijoms, dažniausiai stebimoms tarp dinarų slavų (serbų, kroatų ir bosnių), taip pat Moldovoje ir pietvakarių Ukrainoje. Taip pat pastebimai dažnai aptinkama Albanijoje, šiaurinėje Graikijoje, Bulgarijoje, Slovakijoje, Rytų Ukrainoje, Baltarusijoje ir pietvakarių Rusijoje.
Subkladas I2-M223 būdingas šiaurės vakarų Europai ir pasiekia maksimumą Vokietijoje (Žemutinėje Saksonijoje). Taip pat randama Rumunijoje, Moldovoje ir retai Rytų Europoje.


Šios haplogrupės, kurių dažnis yra nuo 0 iki 95%, yra tarp vokiečių, ispanų, italų, graikų, lenkų, vengrų, prancūzų, albanų, anglų, airių, škotų, portugalų, šveicarų, čekų, slovakų, serbų, kroatų, rumunų, baltarusių. , ukrainiečiai, danai, švedai, norvegai, suomiai, rusai, estai, latviai, lietuviai ir kt.
XX amžiaus 80-aisiais, tiriant genetinio kodo struktūrą, antropologijoje įvyko „tyli revoliucija“. Atsirado nauja mokslo šaka, vadinama paleogenetika arba molekuline paleontologija. Paaiškėjo, kad pačiame žmoguje, o tiksliau jo genotipe, kuris yra visų organizmo genų visuma, galima rasti rūšies evoliucijos istorijos pėdsakų. Pirmą kartą genai pasirodė kaip patikimi istoriniai dokumentai, skirtumas tik tas, kad jie buvo įrašyti ne rašalu, o cheminiuose DNR molekulės komponentuose. Genetikai išmoko pažodžiui išgauti informaciją iš „žemės dulkių“ - suakmenėjusių liekanų, kurios priklausė labai senoms būtybėms. Paleogenetikos gauti duomenys radikaliai pakeitė ankstesnes idėjas apie ankstyvąsias „žmogaus“ evoliucijos stadijas.
Mitochondrinė Ieva – tai molekulinių biologų suteiktas vardas moteriai, kuri buvo paskutinis bendras visų gyvų žmonių protėvis iš motinos pusės. Kadangi mitochondrijų DNR paveldima tik per motinos liniją, visi gyvi žmonės dabar turi tokią DNR iš „Ievos“. Taip pat visų vyrų vyrų Y chromosomos DNR turi būti iš „molekulinio biologinio Adomo“.
Skirtingai nuo branduolinės DNR, kurioje yra didžioji dauguma genų ir lytinio dauginimosi metu vyksta rekombinacija, todėl palikuonys pusę genų gauna iš tėvo, o kitą pusę – iš motinos, vaikas mitochondrijas ir jų DNR gauna tik iš motinos kiaušinėlio. Kadangi mitochondrijų DNR nevyksta rekombinacija, jos pokyčiai gali įvykti tik per retas atsitiktines mutacijas, maždaug kartą per 3000 metų. Palyginus mitochondrijų DNR seką ir laikui bėgant joje atsiradusias mutacijas, galima ne tik nustatyti gyvų žmonių giminystės laipsnį, bet ir apytiksliai apskaičiuoti laiką, reikalingą mutacijų kaupimuisi konkrečioje populiacijoje. žmonių.
Atlikęs lyginamąją mtDNR analizę, 1980 m A. Wilsonas sukonstravo šeimos medį, kuris aiškiai parodė didžiausią mitochondrijų genų diferenciaciją Afrikoje. Be to, visos šešių milijardų šiuolaikinių žmonių populiacijos kilmė yra viena moteris, kažkada gyvenusi Rytų Afrikoje, nes visi tirti mtDNR mėginiai gali būti atsekti iki vienos originalios nukleotidų sekos. A. Wilsonas, radęs vietą, kuri yra žmonijos „lopšys“, pajudėjo toliau. Žinodamas mutacijos greitį, jis sugebėjo nustatyti apytikslį laiką, kada Žemėje pasirodė „Ieva“. „Mitochondrijų laikrodis“ parodė, kad ji gyveno maždaug prieš 200–150 tūkstančių metų (keista, kad „Ieva“ buvo net vyresnė už neandertalietę, kurią „evoliucijos tėvai“ jai atkakliai primetė).
Duomenis iš mtDNR analizės nepriklausomai gavo daugelis kitų tyrėjų. „MtDNR analizė“, – rašo Satoshi Horai,- rodo, kad šiuolaikinis žmogus atsirado maždaug prieš 200 tūkstančių metų Afrikoje, iš kur persikėlė į Euraziją, kur greitai pakeitė Homo erectus ir, tikėtina, visiškai (jei Bigfoot nerastas) neandertalietį. Tuo pačiu metu praktiškai nebuvo mitochondrijų genotipų maišymosi.
1987 metais Rebecca Cann ir kolegos teigė, kad mitochondrijų Ieva galėjo gyventi prieš 140–280 tūkstančių metų. Remiantis naujesniais 2004 m. skaičiavimais, mitochondrinė Ieva gyveno maždaug prieš 140 tūkstančių metų Rytų Afrikoje. Šiuolaikiniai MP ir ME skaičiavimai iš 2009 m. paprastai suteikia Ievos amžiaus intervalą 140 000–230 000 metų, o didžiausia tikimybė yra apie 180 000–200 000 metų.
Ypač įdomios yra įdėtos pastangos L. Cavalli-Sforza bandymas palyginti molekulinės genetikos ir lingvistikos duomenis. Jis parodė, kad genų plitimas stebėtinai gerai koreliuoja su kalbų plitimu. Taigi genetinių tyrimų pagrindu sukonstruotas giminės medis atitinka kalbinį šeimos medį. Taigi genogeografija buvo derinama su etnine geografija.
Dar A. Wilsono gyvavimo metu buvo bandoma analizuoti vyrų Y chromosomą, siekiant atsekti „tėvų liniją“ žmonijos kilmės knygoje. Jo pateikti pirminiai duomenys, gauti prancūzų mokslininko J. Lucotte, taip pat patvirtino afrikietišką „Adamo“ kilmę.
Išsamesnius tyrimus atliko Stanfordo universiteto profesorius P. Underholas, kurie rinko medžiagą analizei beveik visuose pasaulio regionuose. Kaip žinoma, Y chromosoma yra tik vyrų genotipe, todėl per kartas perduodama griežtai iš tėvo sūnui. Ištyrus kelis tūkstančius mėginių, paimtų iš skirtingų tautybių atstovų, paaiškėjo, kad „Adomo“ gimtinė buvo ta pati Rytų Afrika. Tyrėjų teigimu, Homo sapiens vyriškojo atstovo atsiradimo laikas yra apie 150-160 tūkstančių metų. Kai kurie „Ievos“ ir „Adomo“ amžiaus pokyčiai yra metodo klaidos ribose.
Panašius duomenis gavo kita nepriklausoma grupė, vadovaujama Michaelas Hammeris(Arizonos universitetas, JAV). Nurodytas hipotetinio „Adomo“ amžius yra 160–180 tūkst.
Taigi, būtent Afrikos žemyne mūsų protėviai atsirado maždaug prieš 150–200 tūkstančių metų. Maždaug prieš 100 tūkstančių metų jų palikuonys migravo po visą ekumeną, pakeisdami visus kitus ten gyvenusius hominidus, bet, kas svarbu, nesikryžmindami su pastaraisiais. Maždaug prieš 40-60 tūkstančių metų jie pasiekė Europą.
Tačiau tuo paleogenetikų antropologams pateikti netikėtumai nesibaigė. Profesoriui Svante Paabo pavyko išskirti mtDNR iš neandertaliečių slankstelio fragmento, pirmą kartą atrasto 1856 m., gyvenusio maždaug prieš 50 tūkst. Šis darbas išties yra aukščiausia molekulinio genetinio meno viršūnė, o jo rezultatą sunku pervertinti. Kaip parodė lyginamieji šiuolaikinių žmonių ir neandertaliečių mitochondrijų DNR tyrimai, pastarasis visai nėra mūsų protėvis ir net nėra artimas giminaitis. Atlikus lyginamąją „mūsų“ ir „neandertaliečio“ genų analizę, buvo nustatyta, kad skirtumai tarp jų yra tokie dideli, kad šių dviejų rūšių evoliucinės šakos galėjo (ar turėjo) skirtis prieš 600 tūkstančių metų, t.y. laikas, kai pačių rūšių dar nebuvo.
išvadas S. Paabo taip radikaliai keičia idėjas apie antropogenezę, kad iškilo klausimas, ar šiuos rezultatus patikrinti nepriklausoma tyrėjų grupė. Šį kartą dirbau su neandertaliečio kaulo fragmentu Markas Stonekingas, mokslininkas iš grupės A. Wilsonas, taip pat aukščiausias autoritetas paleogenetikos srityje. Atlikęs mtDNR tyrimus iš kito mėginio (prieš 30 tūkst. metų gyvenusio neandertaliečių vaiko palaikų), jis gavo tuos pačius duomenis kaip S. Paabo, visiškai patvirtina jo išvadas. Šiuo klausimu S. Paabo interviu pažymėjo: „Laikėmės griežtų teismo medicinos kriterijų, tarsi ruošėmės teismui pateikti daiktinius įrodymus“.
Po kelerių metų vokiečių mokslininkų grupė taip pat atliko nepriklausomą neandertaliečių mtDNR tyrimą, kuris parodė: „Patvirtinta hipotezė, kad neandertaliečiai yra aklavietės evoliucijos atšaka ir nėra šiuolaikinių žmonių protėviai“.
Paleontologas Christopheris StringerisŠtai kaip jis mato ateitį: „Galbūt esame prie slenksčio sukurti vieningą teoriją, kuri sujungs paleoantropologinius, archeologinius, genetinius ir kalbinius įrodymus Afrikos monogenetinio modelio naudai“.
Iš tiesų, šių mokslų sintezė greičiausiai priartins mus prie mūsų kilmės paslapties supratimo.
Žmogaus mtDNR haplogrupių medis
| Mitochondrijų Ieva | |||||||||||||||||||||||||
| | | |||||||||||||||||||||||||
| | | |||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||
| | | | | | | | | ||||||||||||||||||||||
| iki JT | N1a | ||||||||||||||||||||||||
| | | | | | | |||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||
Žmogaus Y-DNR haplogrupės medis(Y-DNR haplogrupės pagal žmones)
| A1b | A1a-T | |||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A2-T | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2 | A3 | |||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ||||||||||||||||||||||||||||||||||||||||||||||||||
| K | ||||||||||||||||||||||||||||||||||||||||||||||||||
| K(xLT) | ||||||||||||||||||||||||||||||||||||||||||||||||||
Pasvarstykime K- Y-chromosomų haplogrupė su vėlesniais subkladais (L, T, M, NO, P ir S). K-Y chromosomų haplogrupė atsirado prieš 40-50 tūkstančių metų. tikriausiai Vakarų Azijoje.
Naujausiais duomenimis, palikuonys:

Haplogrupės L pasiskirstymas.
Vidutinis induistas
. L haplogrupei būdingas didelis poklasių dažnis ir įvairovė pietvakarių Pakistane, Beludžistane palei pakrantę (28%). Nacionalinė veislė atsirado prieš 24-30 tūkstančių metų.
Papuanų. Nacionalinė veislė atsirado prieš 32-47 tūkstančius metų.
Papuasai Ekari. Nacionalinė veislė atsirado prieš 28-41 tūkstantį metų.
Haplogrupės T pasiskirstymas.
Fulbės yra žmonės, gyvenantys didžiulėje Vakarų Afrikos teritorijoje: nuo Mauritanijos, Gambijos, Senegalo ir Gvinėjos vakaruose iki Kamerūno ir net Sudano rytuose. T haplogrupei būdingas didelis poklasių dažnis ir įvairovė (18%). Nacionalinė veislė atsirado prieš 19-34 tūkstančius metų.
- N haplogrupė.Ši haplogrupė randama Vidurio, Šiaurės Europoje ir visoje Rusijos Europos bei Azijos dalyse. Genetiškai „gryniausi“ atstovai yra jakutai (74%), nencai (74%), udmurtai (68%), suomiai (61%), Uralo kalbų tautos, eskimai. Didžioji dauguma šiuolaikinių šios grupės atstovų priklauso N1 šakai, kurios greičiausiai atsiradimo vieta yra Altajaus regionas, Baikalo sritis, Pietų Sibiras, Mongolija ar Šiaurės Kinija, o laikas yra nuo 20 000 iki 15 000 metų. prieš. Manoma, kad jį per Euraziją atnešė didelė miško Sibiro tautų migracija į vakarus, kurios paskutinė fazė – įsikūrimas Urale, o iš ten – į Volgos sritį ir šiaurės rytų Europą (Rusijos šiaurę, Suomiją, Baltijos šalys) – siejamas su Uralo tautų kalbų plitimu šiame regione.
Nenetai – samojedai, gyvenantys Arkties vandenyno Eurazijos pakrantėje nuo Kolos pusiasalio iki Taimyro. Haplogrupė T turi didžiausią rodiklį (74%) . Tautybė atsirado prieš 15-20 tūkstančių metų.
- Haplogrupė O- Y-chromosomų žmogaus haplogrupė, yra haplogrupės Haplogroup NO palikuonis, pagal įvairias teorijas pirmą kartą atsirado Pietryčių Azijoje arba Rytų Azijoje prieš 28-41 tūkst. būdingas atstovams Mongoloidų rasė. Susijusi su "finougrų" haplogrupe N. Ši haplogrupė yra 80-90% daugumos gyventojų Rytų ir Pietryčių Azijos regione tarp kinų, japonų, filipiniečių, malajų, austroneziečių ir gretimų tautų. veikiamas jų kaip substrato. Šios haplogrupės visiškai nėra Europoje, Vakarų Sibire, Artimuosiuose Rytuose, Afrikoje ir Amerikoje. Tai sudaro apie 21% visos pasaulio žmonių populiacijos.
Haplogrupės O pasiskirstymas.
Vidutinis kinietis - apima visas 56 Kinijoje gyvenančias ir vyriausybės oficialiai pripažintas grupes, tokias kaip mongolai, mandžiūrai, tibetiečiai ir kitos nusistovėjusios etninės grupės, gyvenančios Kinijoje mažiausiai nuo Čing dinastijos laikų (1644–1911). T haplogrupei stebima didelis dažnis ir poklasių įvairovė, didžiausias rodiklis (60-80%). Tautybė atsirado prieš 28-41 tūkstantį metų.
Aukštutinio paleolito eros Rusijos lygumos neoantropinių kaukaziečių fizinės išvaizdos rekonstrukcija:
kairėje – sungirietis (Sungiro, Vladimiro, maždaug prieš 30 tūkst. metų gyvenvietė), dešinėje – Kostenkovietis (Voronežo srities Kostenkų gyvenvietė apie 45 tūkst. metų). Rekonstrukcijos M.M. Gerasimova.
Ši haplogrupė yra padalinta į Q ir R subkladus:
- Q haplogrupė, atsiradimo laikas 15-20 tūkstančių metų prieš Kristų. e., paplitusi tarp kai kurių Sibiro tautų, taip pat tarp vietinių Amerikos tautų ir tam tikru mastu visoje Azijoje. Spėjama, kad šios haplogrupės nešiotojai buvo Sibiro kilmės hunai. Eurazijoje jis randamas trikampyje, kurio viršūnės yra Norvegijoje, Irane ir Mongolijoje. Tačiau iš esmės tarp visų šių tautų tai reta. Europoje ši haplogrupė paplitusi tarp vengrų (2 proc.) ir slovakų (5 proc.). Tačiau ji reikšminga tarp mažųjų Sibiro tautų kumų (95 proc.) ir selkupų (70 proc.). Būdinga ir vietiniams amerikiečiams.
Ketų arba Jenisejų šeima.
Antropologija yra panaši vietinių indėnų ir Okeanijos tautų, pavyzdžiui, polineziečių (Naujoji Zelandija). Tai galimas šios haplogrupės žmogaus įvaizdis 15-20 tūkstančių metų.
- R haplogrupė, haplogrupė R atsirado prieš 30 000–35 000 metų. Skirtingai nuo Q, jis keičiasi ir turi subkladus, o tai rodo jo senumą. Tikėtina, kad tai buvo grupei IJ priklausė kromanjoniečiams (arba daugumai jų), pirmiesiems atstovams
Haplogrupės R pasiskirstymas pogrupiuose R1a (alyvinė) ir R1b (raudona).
- Haplogrupė R2- randama itin retai, daugiausia Irane, Indijoje, Pakistane ir Šiaurės Kaukaze.
- Haplogrupė R1- labiausiai paplitęs haplogrupės R pogrupis. Du pagrindiniai jo subkladai R1a ir R1b (kiti variantai itin reti) yra labiausiai paplitę visoje Europoje ir Vakarų Eurazijoje. Taip yra dėl migracijų po paskutinio ledyno maksimumo. Spėjama, kad haplogrupė R1 galėjo atsirasti prieš 25 000–30 000 metų.
- Haplogrupė R1a, spėjama, kad atsirado Rusijos lygumos pietuose maždaug prieš 10-15 tūkst. Manoma, kad jų pagrindu buvo suformuota slavų etninė grupė. Paplitimo sritis yra nuo Islandijos iki Indijos, modernus haplogrupės centras yra Lenkijoje. Ši haplogrupė tapo kurganų kultūros plitimo žyme, kurią, savo ruožtu, dauguma autoritetingų tyrinėtojų laiko protoindoeuropiečių kultūros šerdimi (Kurgano hipotezė). Genetika parodė, kad pilkapiai skitas skeletuose yra haplogrupė R1a. Plėtra prisidėjo prie haplogrupės R1a migracijos į Iraną ir Indiją, kur jos nešiotojai yra apie 30% vyrų iš aukštesnių kastų. Labiausiai paplitęs Rytų Europoje: tarp luzatų (63 proc.), lenkų (apie 56 proc.), rusų (53 proc.), baltarusių (49 proc.), ukrainiečių (apie 52 proc.), totorių (34 proc.), baškirų. (26%) (tarp Saratovo ir Samaros regionų baškirų iki 48%); ir Vidurinėje Azijoje: tarp chudžando tadžikų (64 proc.), kirgizų (63 proc.), iškašimų (68 proc.). Vidutinis pasiskirstymas Skandinavijos šalyse (23% Islandijoje, 18-22% Švedijoje ir Norvegijoje), Irane (25%?). Tarp Indijos Vakarų Bengalijos ir Utar Pradešo valstijų brahmanų ši haplogrupė pasitaiko atitinkamai 72% ir 67%.
Vidutinis lenkas. Haplogrupėje R1a didžiausias rodiklis (56%).
- Haplogrupė R1b, skirtingi autoriai šios haplogrupės atsiradimą priskiria 16-5 tūkstančiams metų prieš Kristų. e.; Labiausiai paplitęs Europos žemyno vakaruose, ypač didelis jo kalbėtojų procentas yra tarp Vakarų Anglijos tautų, baskų, Pietų Uralo baškirų ir ispanų. Rytų Europoje ir rytuose Haplogrupė R1b yra tarp armėnų, Dagestane. Nešėjas taip pat buvo faraonas Tutanchamonas. Dabartinė R1b koncentracija yra didžiausia pietų Anglijos teritorijose - apie 70%, šiaurės ir vakarų Anglijoje, Velse, Škotijoje, Airijoje - iki 90% ir daugiau, Ispanijoje - 70%, Prancūzijoje - 60%, baskų - 88,1 proc., o ispanai – 70 proc. italai – 40 proc., vokiečiai – 39 proc., norvegai – 25,9 proc.
Vidutinis anglas. Haplogrupėje R1b stebimas didžiausias rodiklis (apie 70%).Tautybė atsirado prieš 10-15 tūkstančių metų.
Norėdami gauti išsamų vaizdą apie šiuolaikinių tautybių atšakų gimimą iki aukščiau aprašyto haplogrupė K Taip pat turėtume pabrėžti kitus, kurie apibrėžia ar stiprina tautiškumą.
Kartu su Kna haplogrupe, po kelių tūkstančių metų haplogrupėIJ (ir jos palikuonys aš Ir J ) . Jo atsiradimo laikas – prieš 38,5 (30,5-46,2) tūkst. Tikėtina, kad pirmiesiems atstovams kromanjoniečiams (arba daugumai jų) priklausė IJ ir P grupės (su R palikuonimis) derinys. šiuolaikinio europietiško tipo žmonės, kuris į Europą atkeliavo maždaug prieš 40 tūkstančių metų, ten sutiko neandertaliečius ir su jais sugyveno 15 tūkstančių metų iki jų išnykimo paskutiniojo ledynmečio metu. Šių kromanjoniečių palikuonys (haplogroup I+R) šiandien sudaro daugumą Europos gyventojų.
I haplogrupės pasiskirstymas.
Haplogrupės J pasiskirstymas.
Vidutinis žiniasklaidos arabas, Saudo Arabija. Haplogrupė J turi didžiausią rodiklį (apie 50%). Tautybė atsirado prieš 20-25 tūkstančius metų.

Gyvenimo prasmė susijusi su klausimu „Dėl ko gyventi“, o ne su klausimu, kaip išlaikyti gyvybę. Žmogaus požiūris...

Grybas yra gyvas organizmas, sudarantis atskirą to paties pavadinimo karalystę. Ilgą laiką jie buvo klasifikuojami kaip augalų karalystės dalis. Tačiau į...

„Ramios“ medžioklės mėgėjams grybų sezonas prasideda vasaros pradžioje ir tęsiasi iki vėlyvo rudens. Ir jie retai...

Aleshnikova, V.I. Profesionalių konsultantų naudojimas. - M.: Infra-M, 1999. - 240 p. 2. Beichas, E....

Apelsinų sultys. Simbolinė apelsinų sulčių reikšmė svajonių knygose yra malonumas ir pagunda. Gana dažnai mes...

Dažniausiai įveikiame visokius gyvenimo kelyje sutiktus sunkumus. Žinoma, tam mes darome...

Šiais metais jūsų globėjas Neptūnas bus jūsų žvaigždyne ir tai geras ženklas, nes jūs...

1993 kas? Kokio gyvūno metai yra 1993 m.? — Pagal kinų horoskopą 1873, 1933, 1993 priklausė Juodųjų metams...

Šviesos bangų ir dalelių dvilypumas reiškia, kad šviesa vienu metu turi nenutrūkstamo...

Biologijos vaidmuo mūsų pasaulyje yra didžiulis. Nors tai nėra vienas iš prioritetinių dalykų, dauguma moksleivių ir...
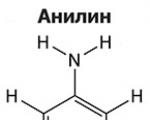
Aminai yra organiniai amoniako dariniai, turintys NH2 amino grupę ir organinį radikalą. Apskritai...
Kaip atsakyti į klausimus B dalyje Antroji socialinių mokslų darbo dalis susideda iš 7 užduočių su trumpu atsakymu....

TORG-15 forma surašoma tuo atveju, kai transportavimo, judėjimo tarp sandėlio ir sandėlio viduje metu, kai...

Mitybos specialistai teigia, kad norėdami geros sveikatos ir lieknos figūros, į savo...

Grybas yra gyvas organizmas, sudarantis atskirą to paties pavadinimo karalystę. Ilgą laiką jie buvo klasifikuojami kaip karalystė...

„Ramios“ medžioklės mėgėjams grybų sezonas prasideda vasaros pradžioje ir tęsiasi iki vėlyvo rudens. Ir jie retai...