Ordem composta Ferramentas e materiais
Ordem toscana. Ordem Toscana, uma das cinco ordens arquitetônicas romanas. O nome está associado à arquitetura etrusca...
COMPLEMENTO(lat. adição de complemento) - um sistema polimolecular de proteínas de soro de leite, um dos fatores mais importantes da imunidade natural. Funções no sangue de humanos, animais de sangue frio e de sangue quente. Contido na linfa e nos fluidos teciduais. Por estar incluído na composição dos imunocomplexos, K. realiza a lise de antígenos celulares sensibilizados por anticorpos, determina a reação de adesão imunológica (ver), participa da opsonização de bactérias, vírus e antígenos corpusculares, acelerando sua fagocitose, e participa de o desenvolvimento de inflamação.
K. foi descrito pela primeira vez sob o nome de “alexin” no final do século XIX. como um fator termolábil inespecífico que determina as propriedades bactericidas do soro sanguíneo fresco (G. Bukhner, 1889). O termo “complemento” foi introduzido por P. Ehrlich (1900), que acreditava que o fator bactericida complementa o efeito citolítico dos anticorpos.
São conhecidas pelo menos 18 proteínas que compõem o sistema K. Estes incluem 9 componentes K, 8 dos quais são proteínas individuais e um é um complexo: 4 proteínas do sistema apropriado, 1 enzima inibidora e 2 enzimas inativadoras.
De acordo com a nomenclatura adotada pela OMS, o sistema K é designado pelo símbolo C, seus componentes individuais por números (C1, C2...C9) e fragmentos de componentes K por letras minúsculas (por exemplo, SZa). A presença de atividade enzimática em um fragmento é indicada por uma linha acima do seu símbolo, e a presença de um centro de ligação com a membrana celular é indicada por um asterisco próximo ao seu símbolo [K. F. Austen et al., 1968].
Os componentes de K. circulam no sangue na forma de precursores, sem se combinarem com anticorpos ou antígenos livres. São descritos dois biol, mecanismos de ativação (ligação) do sistema K. - clássico e chamado. alternativa, ou propriedade [Müller-Eberhard (H. J. Muller-Eberhard), 1975; Vogt (W. Vogt), 1974].
O mecanismo clássico de ativação de K. é realizado com a participação de anticorpos IgG e IgM que fazem parte de complexos imunes, ou imunoglobulinas agregadas de forma inespecífica dessas classes. Quando combinados com antígenos ou como resultado de agregação inespecífica, centros que se ligam a C1, o primeiro componente do sistema K, são formados nas moléculas dessas imunoglobulinas (A. Ya. Kulberg, 1975). Fixado na imunoglobulina C1 inicia uma cadeia de reações nas quais os componentes restantes do sistema K entram sequencialmente.
C1 é um complexo de três subcomponentes (C1q, C1rr e C1s) formados na presença de íons cálcio; C1q é uma proteína semelhante ao colágeno com mol. pesando (massa) 400.000, consistindo em seis subunidades idênticas ligadas de forma não covalente. Cada subunidade contém um centro de reconhecimento para ligação a uma molécula de imunoglobulina. A ligação de C1q à imunoglobulina é acompanhada pelo rearranjo intramolecular de C1q e pela ativação da pró-enzima associada Clr, que atua na proesterase de C1s. A C1s-esterase (C1s) resultante afeta o segundo (C2) e o quarto (C4) componentes do K na fase líquida.
A molécula C4 (peso molecular 208.000) é construída a partir de três cadeias peptídicas - alfa, beta e gama, conectadas por ligações dissulfeto. C1s separa o peptídeo C4a da cadeia alfa, mol. o peso é 8.000, e no fragmento C4b restante da molécula aparece um centro de ligação com a membrana celular sensibilizada por anticorpos. Quando C1s atua sobre C2, mol. cujo peso é 117.000, dois fragmentos são formados - C2b (peso molar 37.000) e C2a (peso molar 80.000). Neste último, é formado um centro de ligação para C4b. O complexo C42 formado na membrana celular é capaz de clivar C3; portanto, é chamada de S3 convertase.
A molécula SZ (peso molecular 180.000) é construída a partir de duas cadeias peptídicas - alfa e beta. Como resultado da clivagem do peptídeo C3 da cadeia alfa pela C3 convertase com mol. pesando 9.000 no fragmento C3b da molécula, um centro de ligação com a membrana celular é formado e um complexo C423 com atividade peptidase em direção a C5 (C5 convertase) é formado na membrana.
Após a clivagem proteolítica de C5, inicia-se a montagem da unidade de ataque à membrana a partir da chamada. componentes terminais do sistema K. A molécula C5 é construída de forma semelhante a S3 a partir de duas cadeias peptídicas a e p, mol. cujo peso é 110.000 e 70.000, respectivamente.A convertase C5 separa o peptídeo C5a da cadeia alfa com um mol. pesando 16.500. O fragmento C5b resultante tem a capacidade de sorver sequencialmente uma molécula de C6 e C7. O complexo C567 absorve uma molécula C8 e seis moléculas C9. No momento da formação, o complexo C5-9 ataca a membrana celular, causando sua destruição. A atividade citolítica do complexo é determinada por C8 e é significativamente aumentada por C9.
Junto com os componentes citolíticos, quando o sistema K é ativado, formam-se os peptídeos fisiologicamente ativos C3a e C5a, chamados anafilatoxinas; eles causam a liberação de histamina pelos mastócitos. contração dos músculos lisos e aumento da permeabilidade vascular, além de servir como fatores quimiotáticos para células polimorfonucleares. A migração dirigida de células polimorfonucleares no local da ativação de K. também é causada pelo complexo trimolecular C567 [Ward (P. Ward), 1975]. Outro peptídeo biologicamente ativo que aparece após a ativação do sistema K é o C3b. Ao se ligar à membrana celular, adquire um segundo centro de ligação estável em relação aos receptores localizados na superfície de diversas células (macrófagos, plaquetas, eritrócitos). Este processo, denominado adesão imunológica, aumenta a fagocitose de células carregadas com K. e partículas corpusculares [S. Ruddy, 1974].
K. também participa do mecanismo de resistência inespecífica a infecções. Nesse caso, o sistema K. é ativado sem a participação de anticorpos por polissacarídeos ou lipopolissacarídeos que fazem parte das paredes celulares de bactérias, leveduras, plantas ou IgA agregada. A ligação de K. ocorre ao longo de uma via alternativa, começando em C3, contornando os estágios de ativação de C1, C4 e C2. Foi demonstrado que a proteína séricapropriadina, o ativador da convertase C3 e vários de seus precursores participam da formação das convertases C3 e C5 da via alternativa. Quando o K é ativado pela via alternativa, assim como pela clássica, forma-se o complexo citolítico C5-9, bem como os peptídeos fisiologicamente ativos C3a e C5a. Este mecanismo provavelmente está subjacente à eliminação inespecífica de vírus e eritrócitos alterados do corpo [L. Pillemer, 1954, 1955].
Todas essas funções dos produtos de reação dos componentes de K. visam a destruição e rápida remoção de informações do corpo. ou agentes estrangeiros. Eles determinam a importância do sistema K como fator de proteção do corpo.
Além de sua função protetora, o sistema K pode contribuir para danos aos próprios tecidos do corpo em diversas doenças com componente autoimune (glomerulonefrite, lúpus eritematoso sistêmico, arterite, miocardite, endocardite). Nesse caso, a ativação do sistema K. é realizada tanto por anticorpos direcionados contra os tecidos quanto por complexos imunes solúveis ou fixos nos tecidos. Os complexos resultantes C423 e C5-9 dos componentes K são fixados em células sensibilizadas e não sensibilizadas por anticorpos, causando destruição de suas membranas. Um papel importante no processo autoimune também pertence aos peptídeos C3a e C5a e ao complexo C567 [N. R. Cooper, 1974; LG Hunsicker, 1974; McCluskey (R. Mc Cluskey), 1975].
O conteúdo de K. é mais frequentemente avaliado por sua atividade hemolítica contra eritrócitos de ovelhas sensibilizados com hemolisina de coelho. O título de K. é expresso em 100 ou 50% de unidades hemolíticas (CH100 ou CH50), ou seja, a quantidade mínima de K., que nas condições experimentais padrão selecionadas lisa, respectivamente, 100 ou 50% dos eritrócitos sensibilizados de maneira ideal. O conteúdo de K. também pode ser avaliado por seu efeito citolítico no sistema sérico linfócito-antilinfócito [Terasaki (R. I. Terasaki), 1964]. K., que não possui atividade lítica, por exemplo. K. cavalo, touro, camundongo, pode ser determinado na reação de aglutinação de eritrócitos sensibilizados carregados com K. com proteína sérica bovina - conglutinina (ver Conglutinação).
Os componentes individuais de K. são titulados em um teste hemolítico usando reagentes especiais, que são preparações de soro fresco de cobaia, desprovidos apenas do componente titulado e contendo os demais componentes em excesso. Os intermediários de hemólise correspondentes também podem ser usados como substratos de titulação. Métodos imunoquímicos e de titulação utilizando anti-soros para componentes puros de K são amplamente utilizados.
O conteúdo de K. nos soros de animais de diversas espécies, de acordo com a titulação hemolítica, varia muito. Seu título mais alto, atingindo 200 CH50 por 1 ml, foi determinado em cobaias. 1 ml de soro humano contém em média 70 e coelho 20 CH50 [R. Audran, 1959, 1960]. Porém, os títulos de K. no teste hemolítico nem sempre correspondem ao seu verdadeiro conteúdo. Assim, K. de algumas espécies não lisa eritrócitos de ovelhas sensibilizados, embora se ligue a eles. A atividade hemolítica de K. de diferentes tipos não é a mesma quando testada em vários sistemas hemolíticos [Boyd (W. S. Boyd), 1969].
Biol, as propriedades de K. de vários tipos são em grande parte determinadas pelo conteúdo de componentes individuais neles. As diferenças de espécies são especialmente pronunciadas no conteúdo de C2 e C4. Esses componentes estão completamente ausentes ou contidos em títulos muito baixos nos soros de cavalos, touros e camundongos, que não possuem atividade lítica. Soros de todos os tipos são caracterizados por um alto teor de C1. O conteúdo dos componentes K. no soro humano é determinado em unidades de peso.
As flutuações individuais no nível e na composição de K. em pessoas saudáveis com idade entre 8 e 35 anos são insignificantes e não dependem do tipo sanguíneo e do fator Rh. Normalmente, as mulheres contêm 10% menos K do que os homens, e em recém-nascidos e mulheres grávidas seu conteúdo é reduzido em média 30% [J. Gumbreitier et al., 1960, 1961]. Houve tendência de aumento dos níveis de K. entre as idades de 35 e 60 anos.
O conteúdo de K. no soro dos pacientes depende da natureza da doença. Na maioria das infecções agudas de etiologia purulenta, assim como na bacteremia estafilocócica, observa-se no período inicial aumento dos títulos de K. Acredita-se que isso esteja associado à ativação de células do sistema reticuloendotelial, em particular macrófagos que sintetizam C2 , C4, C5. Durante o período de eliminação de antígenos com participação de anticorpos, os títulos de K. diminuem e atingem o normal durante a recuperação. Em uma série de doenças que afetam as células do parênquima hepático, por exemplo, cirrose, hepatite, hron, colecistite, a síntese dos inibidores C3, C6, C9 e C1 é perturbada, o que leva a uma diminuição no nível geral de K. Como regra, o nível de K. diminui com condições alérgicas, doenças autoimunes e doenças de complexos imunes devido à ligação de K. circulando no sangue e associado em tecidos com complexos imunes. São descritos casos de deficiência de componentes individuais de K., acompanhados de várias condições patológicas.
O sistema K. está ativo no corpo e em soros recém-isolados. K. é inativado dentro de 2-4 dias quando os soros são armazenados na geladeira (t° 5°), e como resultado do aquecimento dos soros a t° 56° - por 20 minutos. A inativação de K. sob a influência de vários agentes físicos foi descrita. fatores - luz solar, radiação ultravioleta, tremores, sob a influência de produtos químicos. agentes - soluções fracas de ácidos, álcalis, solventes orgânicos, enzimas proteolíticas (L. S. Reznikova, 1967). A atividade de K. permanece por muito tempo em soros liofilizados, quando sulfato de sódio (5%) e ácido bórico (4%) são adicionados aos soros frescos, em soros armazenados a uma temperatura de -40° e abaixo.
A capacidade de K. de ser incluído em complexos imunes é usada para detectar anticorpos e antígenos (ver reação antígeno-anticorpo, reação de fixação do complemento). No entanto, deve-se ter em mente que muitos anti-soros e alguns antígenos se ligam a K. de forma inespecífica. Esse fenômeno, denominado efeito anticomplementar, se expressa na diminuição da atividade hemolítica do K. Pode ser devido à mistura de globulinas agregadas, lipopolissacarídeos ou enzimas proteolíticas nas preparações tituladas, bem como à contaminação bacteriana das preparações (Boyd, 1969). O aumento da capacidade dos anticorpos de alguns indivíduos da mesma espécie para fixação inespecífica de K. é chamado de desviabilidade, e os anticorpos que possuem essa propriedade são chamados de desviáveis.
O estudo do processo de ativação de K., esclarecimento do biol, propriedades dos produtos de ativação dos componentes de K., nível de K. em condições normais e em diversas doenças permite compreender sua função protetora e seu papel no tecido dano. Este conhecimento é necessário, em particular, para o desenvolvimento de métodos com base científica para a prevenção e tratamento de doenças causadas pela activação do sistema K.
A determinação dos títulos de K. em diversas doenças ao longo do tempo é de importância prática, pois é um indicador do imunol, do estado do corpo, da eficácia do tratamento. atividades e tem significado prognóstico.
Bibliografia: Boyd W. Fundamentos de imunologia, trad. do inglês, pág. 346, M., 1969; Inflamação, imunidade e hipersensibilidade, ed. G. 3. Moveta, pista. do inglês, pág. 422, M., 1975, bibliografia; Kulberg A. Ya. Imunoglobulinas como reguladores biológicos, p. 106, M., 1975, bibliografia; Cabot E. iMeyer M, Imunoquímica Experimental, trad. do inglês, pág. 140, M., 1968, bibliografia; P e z n e para a ilha e L. S. Complemento e seu significado nas reações imunológicas, M., 1967, bibliogr.; Austen K. F. a. ó. Nomenclatura de complemento, Boi. Wld Hlth Org., v. 39, pág. 935, 1968; Col ten HR Biossíntese de complemento, Advanc. Imunol., v. 22, pág. 67, 1976, bibliografia; Imunologia abrangente, ed. por N. K. Day a. R. A. Bom, v. 2, Nova York, 1977; Complemento Muller-Eberhard HJ, Ann Rev. Bioquímica, v. 44, pág. 697, 1975, bibliografia; Yogt W. Ativação, atividades e produtos farmacologicamente ativos do complemento, Pharmacol. Rev., v. 26, pág. 125, 1974, bibliografia.
IA Tarkhanova.
Estrutura do sistema complemento:
1. proteínas – ativadores (componentes iniciais) – C1qrs, C2, C3, C4.
2. proteínas da sequência final (componentes terminais) – C5, C6, C7, C8, C9.
3. fatores da via alternativa de ativação – D i, B i, P.
4. inibidores e ativadores de proteínas:
· possuir atividade enzimática – fator I, C3b – inativador, carboxipeptidases.
· Proteínas de ligação e concorrentes - C4BP (proteína de ligação a C4), vitronectina ou proteína S, C1 - INH (C1 - inibidor), fator H, DAF (CD55 - fator potenciador de putrefação), CD59, MCP (CD46).
C2b é inativado por C4BP e DAF, C3b é inativado pelo fator H CR1 b.
5. receptores para proteínas do complemento. Designado CR - receptor de complemento.
CR3(CD11b, CD18)
CR4(CD11c, CD18)
O sistema complemento consiste em cerca de quarenta proteínas plasmáticas funcionalmente relacionadas: componentes, fatores proteicos reguladores e receptores. São capazes de ativação sequencial em forma de cascata com efeito citotóxico final inespecífico contra bactérias, fungos, vírus, células tumorais, células transplantadas e outros.
O complemento está incluído em Megassistemas Hagemann, juntamente com o sistema de coagulação sanguínea, sistema fibrinolítico e cinina-calicreína.
Fragmentos– são produtos da quebra de componentes: C3a, C3b, etc. Se o fragmento estiver inativado, adicione a letra I: C3ai; proteínas com atividade enzimática são indicadas por uma linha acima da letra D -, C4α, C4β são cadeias individuais em moléculas peptídicas.
A biossíntese de proteínas do complemento ocorre no fígado, epitélio do intestino delgado, medula óssea, baço e macrófagos.
Síntese e consumo, ativação e inibição estão em equilíbrio lábil e são fortemente regulados. Em alguns casos, as cadeias polipeptídicas de um componente C1, C8 são sintetizadas separadamente e montadas imediatamente antes da secreção como uma arma binária. A síntese do complemento no período embrionário inicia-se a partir da 6ª semana de desenvolvimento, e na 10ª semana já é registrada atividade citotóxica.
Mecanismos de ativação do complemento:
1. via clássica – dependente de complexos imunes
2. lectina – perto do clássico
3. alternativa – dependente do fator P
4. protease – próxima da alternativa, mas não depende do fator P.
Caminho de ativação clássico.
É desencadeada pelo complexo antígeno + anticorpo (complexo imune - RI), ou seja, requer a presença de um processo imunológico específico no momento. Pode ser desencadeada pelo complexo antígeno + proteína C reativa, ou pela lectina ligadora de manose (via da lectina) - MBL. Ao se fixar na superfície da célula alvo (*: bactéria), o anticorpo inicialmente altera o que deveria ser posteriormente destruído pelo complemento. A ligação de um anticorpo com um antígeno leva a uma mudança na configuração espacial do anticorpo, o que possibilita maior fixação de C1q nele.
No entanto, para uma fixação forte, são necessárias 1 molécula de IgM ou 2 moléculas de IgG. Outras classes de Ig não ativam o complemento.
O componente do complemento C1 consiste em três componentes: C1q, C1r, C1s.
C1q é um monômero, C1r é um dímero (pré-protease C1s), C1s é um dímero (pré-protease para C2 e C4).
Externamente, a molécula C1qrs tem o formato de uma tulipa.
Efeitos fisiológicos e patológicos de fragmentos de complemento ativados.
C2a é uma substância semelhante à cinina que irrita os receptores da dor.
C4a – causa dilatação grave dos capilares e inchaço grave dos tecidos
C3b – estimulação de adesão e opsonização
C3a, C5a são anafilotoxinas 1 e 2, respectivamente, estimulam a quimiotaxia das células até o local da inflamação, causando espasmo da musculatura lisa, aumento da permeabilidade capilar e desgranulação dos mastócitos (por exemplo, devido à histamina). C5a é um fragmento mais agressivo que C3a. C5a tem potencial para citólise (são formados muitos radicais dependentes de oxigênio).
C5b6789...9 é uma substância em forma de cilindro oco com centros de ligação lipídica; é um complexo de ataque à membrana. Um cilindro com furo de diâmetro de 10 nm, que, devido aos centros de ligação lipídica, é capaz de se fixar na membrana da célula alvo, perfurando-a, fazendo com que o conteúdo da célula flua (mais precisamente, íons de sódio e água entram na célula).
Funções básicas do complemento.
1. microbicida – dirigido contra bactérias, vírus, fungos
2. citolítico - contra células tumorais de transplantes, em caso de patologia - contra as próprias (em caso de defeitos regulatórios e picadas de insetos ou répteis).
3. participação na inflamação – C2a, C3a, C4a, C5a
4. ativação da reação fagocítica: quimiotaxia, opsonização, adesão e absorção. O fragmento principal é C3b, mas a quimiotaxia também é estimulada por C3a e C5a.
5. interação com outras partes do megassistema Hageman: fator de coagulação 12 - capaz de ativar o sistema complemento por via alternativa.
Calicreína, plasmina, trombina – ativam C3, C4, C5, B.
6. participação na regulação da resposta imune
7. participação em reações alérgicas – anafilotoxinas.
Complementoé um sistema que consiste nas próprias proteínas do complemento, receptores de membrana para complemento, reguladores plasmáticos e de membrana da atividade do complemento.
As próprias proteínas do complemento são uma série de glicoproteínas e fatores proteicos no plasma sanguíneo, incluindo 9 componentes diferentes. Eles formam uma cascata molecular multienzimática, na qual o produto de uma reação é o substrato para a próxima. Neste caso, ocorre um aumento gradual do potencial lítico e um estímulo inicial inicialmente fraco leva a um poderoso efeito antimicrobiano final.
Existem 4 tipos de receptores para componentes do complemento (receptor do complemento, CR - I, II, III, IV). O primeiro tipo de receptor (CR) é encontrado na superfície das células apresentadoras de antígenos e dos glóbulos vermelhos. Ele medeia a captura de um patógeno ao qual as opsoninas C3b e C4b estão ligadas. A ligação dos complexos imunes pelos eritrócitos garante seu transporte para o fígado e baço, onde estão contidos os macrófagos. O segundo tipo de receptor (CR II) é expresso em linfócitos B e células dendríticas foliculares. Participa da fixação por essas células de complexos imunes nos centros germinativos dos folículos dos gânglios linfáticos, causando maior hipermutagênese somática dos receptores de imunoglobulina dos linfócitos B e a formação de células B de memória. CR III e CR IV por sua natureza pertencem a β 2 -integrinas (moléculas de adesão) e são específicas para iC3b (inativado sob a influência do fator H) e C3d. Esses receptores são encontrados predominantemente nos fagócitos e desempenham uma função dupla. Primeiramente, promovem a migração de fagócitos para o local da inflamação, pois podem interagir com as moléculas de membrana de adesão ICAM-1 e ICAM-2, cuja expressão nas células teciduais é um dos marcos para o movimento direcionado. Em segundo lugar, os fagócitos, penetrando na lesão graças ao CR III CR IV, reconhecem os componentes do complemento, o que promove a fagocitose do patógeno marcado com opsonina.
Juntamente com numerosos componentes do complemento, circulam no plasma sanguíneo proteínas com propriedades antagônicas, que limitam a ativação do sistema complemento durante a neutralização do patógeno. Um dos mais importantes é o inibidor do primeiro componente (inibidor C1), cuja deficiência aumenta o risco de desenvolver angioedema hereditário. O chamado fator H garante a inativação de C3b, promovendo sua posterior clivagem nos fragmentos C3c e C3d, e o fator I destrói C3b e C4b.
Como você pode ver, 2 fatores plasmáticos neutralizam o C3b ao mesmo tempo. Isso é necessário para o correto funcionamento da via alternativa, uma vez que o excesso desse fragmento causa hiperativação irracional do complemento, causando graves autodanos. O início da cascata ocorre justamente devido ao C3b, formado durante a hidrólise espontânea do C3. Ressalta-se que a hidrólise espontânea é sempre limitada, o que evita uma possível hiperativação do sistema. Ao mesmo tempo, sob a ação da C3 convertase, o fragmento C3b é formado em quantidade suficiente para iniciar uma nova cascata, cujo desdobramento libera uma porção adicional de C3b. Devido a este feedback positivo, o complemento aumenta o seu potencial lítico enquanto a quantidade de patógeno permanece inalterada. No entanto, se o processo descrito não for devidamente controlado, é bem possível a hiperativação irracional do complemento ao longo da via alternativa e, como resultado, danos aos próprios tecidos. Além disso, o mecanismo alternativo também pode ser potencializado pelo C3b, liberado como resultado da via de ativação clássica que ocorre em paralelo. Portanto, para o correto funcionamento de todo o sistema, é necessária a inativação adequada do C3b resultante.
Como o complemento realiza o reconhecimento de padrões e as estruturas de membrana das próprias células normalmente sofrem alterações dinâmicas, existe um perigo potencial de autoagressão mediada pelo complemento. Para evitá-lo, proteínas protetoras são “incorporadas” nas membranas das próprias células, inativando a cascata do complemento. Estamos falando de um fator que acelera a cárie (inglês: Decay accelerating factor, DAF), que é encontrado nas células sanguíneas, células epiteliais e células endoteliais. Aumenta o catabolismo das principais enzimas da cascata - C3 e C5 convertases. As proteínas protetoras da membrana também incluem a proteína cofator de membrana (MCP), que é um cofator na proteólise de C3b e C4b usando o fator 1.
O papel do sistema complemento é: Matéria do site
Sistema complementoé um complexo complexo de globulinas séricas. Este sistema em cascata de enzimas proteolíticas destina-se à proteção humoral do corpo contra a ação de agentes estranhos e está envolvido na implementação da resposta imunológica do corpo. As proteínas do sistema complemento fornecem uma resposta rápida e eficaz a um sinal inicialmente fraco e trazem consequências funcionais. Os componentes do sistema complemento são geralmente designados por letras latinas.
Existem dois mecanismos para ativar o sistema complemento:
clássico;
alternativa.
Esses mecanismos são conectados ao nível do 5º componente e depois procedem da mesma forma.
A maneira clássica.
O mecanismo desencadeante é a formação de um complexo antígeno-anticorpo (AG-AT) na superfície da célula-alvo. Ao mesmo tempo, ocorrem alterações conformacionais na molécula de imunoglobulina (é designada: Ig ou AT). Como resultado dessas alterações, a Ig adquire a capacidade de se ligar ao componente C 1 q do complemento. C 1 r e C 1 s se unem a eles, e todo esse complexo sofre um rearranjo conformacional e se transforma em uma C 1 esterase, que atua sobre C 4, C 4 a é clivado e C 4 b faz parte do complexo. Então C2 se une ao complexo, formando um novo substrato para ação de C1s, C2b é clivado e C2a passa a fazer parte do complexo.
O complexo resultante é denominado “C 3 -convertase”, e sob sua ação o peptídeo C 3 a é clivado, e C 3 b faz parte do complexo, que agora é denominado “C 5 -convertase”. A C5 convertase atua em C5, cliva C5a dele e C5b faz parte do complexo.
Depois disso, C 6, C 7 e C 8 são sequencialmente associados a C 5 b. Como resultado, forma-se um complexo capaz de ligar 2 moléculas C9.
Se esse processo ocorrer na superfície da célula alvo, então os componentes do complexo C 5 b-C 9 formam um complexo de ataque à membrana, que forma canais transmembrana na superfície da célula alvo que são completamente permeáveis a eletrólitos e água. A célula alvo morre.
Os subprodutos (menores) do processo C 3 a e C 5 a possuem propriedades de anafilotoxinas.
Regulação da via clássica.
A maioria dos componentes está ativa apenas como parte do complexo. Suas formas ativas podem existir por um período muito curto. Se durante esse período não atenderem ao próximo componente, as formas ativas perdem o contato com o complexo e tornam-se inativas. Se a concentração de qualquer componente estiver abaixo do limite (crítico), o funcionamento do sistema complemento não levará a consequências fisiológicas.
Os inibidores endógenos da proteinase também participam da regulação do sistema complemento. O mais eficaz deles é o inibidor C 1.
Caminho alternativo.
A diferença entre a via alternativa e a clássica é que ela não requer a formação de complexos imunes para ser desencadeada.
O mecanismo desencadeante da via alternativa é a formação de C 3 b a partir de C 3 sob a influência de algum fator desencadeante: por exemplo, polissacarídeos da parede celular bacteriana.
C3b forma um complexo com o fator “B” (C 3 bB), que é exposto à protease D (sempre ativa no plasma sanguíneo!). Como resultado, “Ba” é clivado e forma-se o complexo C3bBb, que tem atividade proteolítica em relação a C5 - ele cliva C5a dele.
Depois disso, as reações prosseguem da mesma forma que na forma clássica.
O substrato para C 3 b também é C 3, como resultado do qual uma quantidade ainda maior de C 3 b é formada - um feedback positivo é observado. Portanto, mesmo pequenas quantidades de C 3 bBb são suficientes para obter cada vez mais sua forma ativa (amplificação do sinal inicialmente fraco).
A via alternativa normalmente funciona sempre e de forma muito ativa, o que proporciona uma resposta rápida e inespecífica à introdução de células estranhas.
Inibidores específicos participam da regulação do sistema complemento, que regulam a velocidade das enzimas das principais reações.
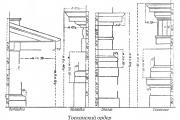
Ordem toscana. Ordem Toscana, uma das cinco ordens arquitetônicas romanas. O nome está associado à arquitetura etrusca...

LIBRETO. Se você ama música (o que sem dúvida ama, já que precisava deste livro), então provavelmente já teve...

Religião: paganismo Nascimento: 942 (0942) Morte: Marcha no Dnieper Família: Rurikovich Pai: Igor...

Masaccio (na verdade Tommaso di Giovanni di Simone Cassai (Guidi), Tommaso di ser Giovanni di Guidi; 21 de dezembro de 1401...

(Publicado em Tel Aviv em 1958. Transmitido com pequenas abreviaturas). Judeus! Amem-se, ajudem-se...

Pegue uma concha ou panela e despeje leite e creme nela. Adicione o açúcar baunilhado (açúcar com sementes de baunilha). Em separado...

Olá pessoal! Hoje temos diante de nós uma receita muito saborosa e simples - salsichas em massa no forno. Você sente...

O abacate é uma das frutas mais saudáveis. Este é um verdadeiro depósito de vitaminas e nutrientes. E ele...

O porco assado é um prato ideal, saboroso e satisfatório, que pode alimentar toda a família. Senhoras...

Este bolo é muito fácil de fazer e tem gosto de clássico Napoleão. Na hora de preparar o bolo existe...

Os representantes desses signos do zodíaco nem sempre se tornam pessoas próximas, mas são agradáveis e interessantes uns com os outros....

A compatibilidade entre um homem Galo e uma mulher Porca baseia-se no entendimento mútuo entre os parceiros. Eles têm personalidades diferentes e...

vários conceitos e fórmulas básicas. Todas as substâncias têm massa, densidade e volume diferentes. Um pedaço de metal...

Se vamos falar sobre igualdade de gênero, então será com alguém como a mulher de Áries. Esta senhora emancipada é uma pessoa brilhante...

LIBRETO. Se você gosta de música (o que sem dúvida gosta, já que precisava deste livro), então provavelmente...

Religião: paganismo Nascimento: 942 (0942) Morte: Marcha no Dnieper Família: Rurikovich Pai:...