Що можна і не можна на Різдвяний піст?
У 2018 році Різдвяний пост розпочнеться 28 листопада. У цей період православні віруючі готуються до зустрічі Різдва.
Тема лекції: "Нейролептики, транквілізатори, седативні засоби".
Нейролептики
В даний час у групу нейролептиків об'єднано близько 500 препаратів.
![]() Класифікація
Класифікація
А. «Типові» Б. «Атипові»
нейролептики: нейролептики:
-аміназин - азалептин
Трифтазін
Галоперидол
Дроперидол
Родоначальником нейролептиків є аміназин, який був синтезований в 1950 р. Шарпентьє (Франція), а вивчений Курвуазьє.
| Препарати | Механізм дії | Застосування |
| Аміназин (Aminazinum) ін.0,025; 0,05; 0,1; амп. 2,5% по 1 мл, 2 мл, 5 мл., внутрішньом'язово і внутрішньовенно | 1. Антипсихотична дія (усунення абсурду, галюцинацій) проявляється через 1-2 тижні. після початку лікування. 2.Седативну дію (усунення страху, тривоги, занепокоєння) проявляється через 15 хв. після внутрішньом'язового введення. 3. Протиблювотна дія (усуває та попереджає блювання та гикавку центрального походження). 4. Потенційна дія. 5. Гіпотензивна дія (АТ) 6. Гіпотермічна дія (t) 7. Знижує тонус скелетної мускулатури | Психози (шизофренія, епілепсія, маніакально-депресивний психоз, алкогольний психоз-біла гарячка). Психози, неврози (неврастенія, істерія, невроз нав'язливих станів). Неприборкані блювання вагітних, травми, пухлини мозку, променева хвороба, блювання, викликані лікуванням протипухлинними препаратами. Підсилює дію засобів для наркозу, снодійних, анальгетиків та ін Гіпертонічний криз (рідко). У складі літичної суміші при гіпертермічному синдромі (рідко). |
Побічна дія: сонливість, млявість, при тривалому застосуванні можлива депресія, ортостатичний колапс, ураження печінки, порушення кровотворення, алергічні реакції, паркінсонізм, диспепсичні розлади. Місцево: розвиток дерматитів, при внутрішньом'язовому введенні – хворобливі інфільтрати, при внутрішньовенному введенні – тромбофлебіти.
| Трифтазин (Triftazinum), таб., р-р в ампулах; в/м. Галоперидол (Haloperidolum); таб., розчин у фл, по 10мл (всередину), розчин в ампулах; в/м та в/в Дроперідол (Droperidolum); 0,25% розчин в амп. по 2мл та 5 мл, у фл.по 5 мл; під/до, в/м, в/в. | 1.Антипсихотична дія 2. Протиблювота дія виражена сильніше, ніж у аміназину. 3. Інші властивості слабо виражені, або відсутні. 1.Антипсихотическое дію, галюцинації знімає швидше марення (перевищує аміназин в 50 разів). 2. Седативна дія 3. Протиблювотна дія (перевищує аміназин у 50 разів). 4. Потенційна дія. 5. Протисудомна дія. Інші ефекти, властиві аміназину, виражені слабо. 1. Антипсихотична дія, 2. Седативна дія 3. Протиблювота дія 4. Потенціювальна дія, наприклад фентаніл + дроперидол = таламонал 5. Гіпотензивна дія. Дія розвивається через 5-15 хв., Триває 3-5 годин. | Див.аміназин -//- -//- Див.аміназин Блювота різного походження. Див.аміназин -//- Див.аміназин В анестезіології для аналгезії при підготовці до хірургічних втручань і після них, при підготовці до інструментальних досліджень, при травмах, інфаркті міокарда. Гіпертонічний криз | ||
| Побічна дія:депресія, явище паркінсонізму, гіпотензія, гноблення дихання. | ||||
| Азалептін(Asaleptinum); таб.0,025 та 0,1; амп.2,5% – 2мл; в/м | 1.Антипсихотическое дію виражено сильно 2. Седативне і снодійне дію. 3. Підсилює дію снодійних та анальгетиків. 4. Розслаблює кістякові м'язи. Інші ефекти, властиві аміназину не виражені | Див.аміназин -//- -//- -//- | ||
Режим дозування встановлюється індивідуально, починаючи з невеликих доз, які поступово збільшують. Добову дозу можна застосовувати одноразово перед сном або 2-3 рази на день після їди.
Після досягнення терапевтичного ефекту дозу знижують та переходять на підтримуючий курс.
Побічна дія: сонливість, біль голови, м'язова слабкість, тахікардія, гіпотензія, сухість у роті, порушення акомодації, пітливість, підвищення маси тіла, зниження потенції, пригнічення крові.
Явище паркінсонізму не відзначається.
Протипоказання: вагітність (перші 3 місяці), період лактації, дітям до 5 років, глаукома, міастенія, пригнічення крові, водіння транспорту тощо, епілепсія, алкогольний психоз.
Транквілізатори
I.Виробні II. «Денні» транквілізатори
бензодіазепіну - рудотель
Феназепам - грандаксин
- сібазон (седуксен,
діазепам,
реланіум)
- Нозепам (тазепам)
Алзолам
Побічна дія: сонливість, головний біль, запаморочення, атаксія (хиткість ходи), алергічні реакції, порушення менструального циклу, зниження потенції, у великих дозах можлива амнезія, при тривалому прийомі (до 6 місяців) виникає звикання та пристрасть, синдром відміни.
Протипоказання: ураження печінки, нирок, міастенія, у процесі роботи, що потребує швидкої реакції та координації рухів, заборонено поєднувати з алкоголем, перші 3 міс. вагітності.
«Денні» транквілізатори не мають снодійної дії, не викликають міорелаксації.
Побічні дії грандаксину: алергічні реакції; підвищення збудливості.
Протипоказаний при вагітності.
Седативні засоби
Препарати групи регулюють процеси гальмування та збудження у корі головного мозку.
Існують також інші види конвергенції збудження на нейроні, характерні тільки для кори великого мозку. Це пов'язано з тим, що будь-які периферичні збудження, перш ніж досягти кори великого мозку, зазнають дисперсії по численних підкіркових утворах Г. м., а потім у вигляді висхідних потоків збудженні різної модальності прямують до коркових нейронів. Широка конвергенція збудження різних модальностей дає великі можливості для подальшого кінцевого ефекту взаємодії збуджень. Результат взаємодії різних збуджень на окремому нейроні визначає рівень його подальшої участі у формуванні адекватного поведінкового акта організму.
Результатом такої взаємодії можуть бути явища торування, викриття, гальмування та оклюзії. Проторення полягає у зменшенні часу синаптичної затримки та передачі збудження за рахунок тимчасової сумації імпульсів, що йдуть за аксоном. Ефект полегшення проявляється тоді, коли серія імпульсів збудження викликає в синаптичному полі нейрона стан підпорогового збудження, який сам по собі недостатній для появи на постсинаптичній мембрані потенціалу дії, але за наявності наступної імпульсації, що приходить по будь-яким іншим аксонам і досягає того ж самого синаптичного поля стає пороговим і в нейроні можливе виникнення збудження. У разі одночасного приходу різних аферентних збуджень до синаптичних полів кількох нейронів знижується загальна кількість збуджених клітин (оклюзія), що проявляється зниженням функціональної активності виконавчого органу.
Поодиноке велике аферентне закінчення контактує з великою кількістю дендритів окремих нейронів. Така ультраструктурна організація є основою для широкої дивергенції імпульсів збудження, що призводить до іррадіації збудження в межах будь-якої структури. Іррадіація є спрямованою, коли збудженням охоплюється певна група нейронів. Об'єднання на одному нейроні синаптичних входів від багатьох сусідніх клітин створює умови для мультиплікації (множення) імпульсів збудження на аксоні, а нейронна пастка забезпечує пролонгування збуджень у Р. м. Подібні функціональні зв'язки можуть сприяти тривалій роботі ефекторних нейронів при малій кількості Г, що приходять в центри. м. аферентних імпульсів.
Генералізовані активуючі впливи здійснюються також гіпоталамусом та лімбічними структурами. Виникаючі в клітинах гіпоталамуса збудження за рахунок його широких зв'язків поширюються на інші структури Р. м. Найбільш виражені висхідні активуючі впливи гіпоталамуса спрямовані на передні відділи кори великого мозку, захоплюють лімбічні утворення ядра таламуса. При посиленні збудження центрів гіпоталамуса активуються також ретикулярні структури стовбура Г. м. Лімбічні структури, до яких належать нюховий мозок, парагиппокампальна звивина, скроневі та лобові відділи кори (див. Лімбічна система ), за рахунок внутрішніх взаємозв'язків та широкого впливу на інші утворення Р. м. також сприяють його генералізованому збудженню. Активуючі впливи є фізіологічною основою виникнення мотиваційного порушення головного мозку. При появі внутрішніх потреб організму відбувається збудження мотивації генних центрів гіпоталамуса, лімбічних і ретикулярних утворень, які за рахунок своїх активуючих впливів організують процеси в Р. м., що призводить до формування цілеспрямованої поведінки. Поряд з активуючими висхідними впливами Г. м. існують низхідні, головним чином кортикофугальні, впливи на підкіркові структури. Взаємодія висхідних та низхідних впливів зумовлює двосторонній зв'язок між структурами Г. м., особливо виражену між корою великого мозку та підкірковими утвореннями. Подібна реверберація збуджень може сприяти збереженню вогнищ тривалого збудження, що лежить в основі короткочасних механізмів. пам'яті чи тривалих емоційних станів.
Функціональні зв'язки між різними відділами Р. м., з одного боку, визначають його функції як єдиного цілого, з іншого - відбивають функціональну роль окремих його структур. У загальному вигляді основна функція Р. м. полягає у регуляції функцій цілісного організму. Однак при аналізі кожної конкретної функції можна виявити і конкретні мозкові структури, що регулюють її більшою мірою. Регуляція вегетативних функцій організму зрештою спрямовано підтримку постійності його внутрішнього середовища. Постійність внутрішнього середовища, або гомеостаз, характеризується безліччю показників - гемодинамічним та осмотичним тиском крові, її температурою, рН, кількістю формених елементів, концентрацією цукру, деяких інших речовин та ін. Гомеостаз забезпечують функціональні системи організму, що динамічно організуються за принципом саморегуляції (див. Саморегуляція фізіологічних функцій ). Кожна функціональна система вибірково поєднує різні структури головного мозку, які у взаємодії із залозами внутрішньої секреції здійснюють нейрогуморальне регулювання функцій. Важлива роль цьому регуляції належить гіпоталамо-гіпофізарної системи, центром якої є гіпоталамус. У ньому знаходяться центри голоду, насичення, спраги, терморегуляції, сну та неспання. Отримуючи аферентні потоки збуджень від інтероцепторів (осморецепторів, терморецепторів, хеморецепторів), ядра гіпоталамуса, інтегруючи їх, адресують збудження за функціональними зв'язками до еферентних нейронів парасимпатичного та симпатичного відділів вегетативної нервової системи. Відповідно до зміненого параметра внутрішнього середовища організму здійснюється регулюючий вплив вегетативної нервової системи на відповідний орган (наприклад, нирки, шлунково-кишковий тракт, легені, серце). Крім того, центри гіпоталамуса, так само як і центри лімбічної системи, за рахунок функціональних міжструктурних взаємодій у Г. м. формують відповідний мотиваційний стан організму (наприклад, стан голоду, спраги, статевого потягу), на основі чого організується поведінка людини.
Життєво важливі центри регуляції вегетативних функцій (судинний і дихальний) знаходяться в структурах довгастого мозку. Довгастим мозком здійснюється також регуляція таких найпростіших реакцій, як ссання, ковтання, жування, блювання, чхання, моргання та ін. У регуляції вегетативних функцій бере участь мозок, який впливає на діяльність внутрішніх органів через симпатичний відділ вегетативної нервової системи. Вищим центром регуляції вегетативних функцій є лімбічна система, яка іноді називається вісцеральним мозком. Від лімбічної системи імпульси збудження направляються насамперед до вегетативних центрів гіпоталамуса, через нього до гіпофіза та ядра симпатичного та парасимпатичного відділів вегетативної нервової системи. Завдяки своїм зв'язкам із базальними гангліями, передніми відділами таламуса та ретикулярною формацією лімбічна система може впливати на функціональний стан скелетних м'язів. Деякі області кори великого мозку, особливо лобові та тім'яні відділи, беруть участь у регуляції вегетативних функцій. Роздратування цих областей викликає зміну серцевої діяльності, рівня артеріального тиску та ритму дихання, слиновиділення, руху кишечника, блювання.
Поруч із регуляцією функцій окремих органів Р. м. надає трофічні регуляторні впливу різні клітини і тканини. Центральні механізми регулювання трофіки здійснюються подібно до механізмів регуляції функцій, але опосередковуються переважно через симпатичний відділ нервової системи (див. Вегетативна нервова система ). Симпатичні впливи на клітини, особливо на скелетні м'язи, завжди адаптаційно-трофічні і здійснюються за рахунок медіатора норадреналіну, що виділяється. Адаптаційно-трофічний вплив симпатичної нервової системи може бути і непрямим за рахунок виділення норадреналіну в рідкі середовища організму (кров, цереброспінальну рідину, лімфу) або здійснюватись через гіпоталамус та залози внутрішньої секреції. Більшість дослідників розглядають будь-який вплив нервової системи як трофічний, який здійснюється у безімпульсній формі та має схожість з процесами нейросекреції. Такі речовини, що утворюються в нервових клітинах, як медіатори, пептиди, амінокислоти, ферменти та ін., доставляються до виконавчих органів за допомогою аксонального транспорту і впливають на їх обмін.
Регулюючи рухові функції, Р. м. забезпечує два основні процеси: створення та перерозподіл тонусу скелетних м'язів для збереження пози та координацію послідовності та сили скорочення м'язів для організації руху. Підтримка пози і координація цих процесів з організацією цілеспрямованого руху здійснюються головним чином стовбуровими структурами Р. м., тоді як сама організація та виконання цілеспрямованого рухового акта вимагають участі вищеутворених утворень Р. м., включаючи кору великого мозку. Регуляція м'язового тонусу та формування пози забезпечуються комплексом настановних реакцій, які поділяються на статичні та статокінетичні. Статичні реакції сприяють збереженню рівноваги тіла людини у просторі за зміни становища її окремих частин (голови, рук, ніг). Статокінетичні реакції пов'язані з перерозподілом тонусу скелетних м'язів, що забезпечує збереження рівноваги тіла людини при кутових та лінійних прискореннях активного чи пасивного переміщення їх у просторі. Вестибулярні ядра довгастого мозку є першим рівнем в Р. м., де відбувається обробка інформації, що надходить від рецепторів лабіринту про рух або зміну положення тіла в просторі. Нейрони цих ядер одержують аферентні потоки також від пропріоцепторів м'язів і сухожиль, що виникають при зміні положення частин тіла в просторі, а також впливу з боку інших структур Р. м. (мозочка, ретикулярної формації, базальних ядер, моторної області кори великого мозку). Східні регулюючі впливи Р. м. на скелетні м'язи здійснюються через сегментні механізми спинного мозку.
Вищим рівнем регуляції рухів є кора великого мозку, базальні ядра та мозок. До рухових областей кори великого мозку відносяться первинна та вторинна моторні області та премоторна область. Цитоархітектоніка первинної рухової області характеризується сукупністю вертикальних колонок нейронів, кожна з яких забезпечує збудження або гальмування однієї групи мотонейронів, що іннервують окрему м'язи. Т.ч., в соматотопічній організації рухової області представлені м'язи всіх частин тіла людини, причому переважно м'язи пальців рук, губ і язика, меншою - м'язи тулуба і нижніх кінцівок. Від рухової кори починається пірамідний, або кортикоспінальний шлях прямої регуляції активності мотонейронів спинного мозку для здійснення тонких рухів (наприклад, артикуляція, втягування нитки в голку). Багато загальних рухових актів (наприклад, ходьба, біг, стрибки) відбуваються без участі пірамідної системи, але з обов'язковою участю екстрапірамідної системи. Центральне місце серед структур екстрапірамідної системи посідають базальні ядра. За допомогою цих структур досягаються плавність рухів та встановлення вихідної пози для їх здійснення. Більшість структур екстрапірамідної системи немає прямих виходів до мотонейронів спинного мозку, а опосередковує свій вплив них через ретикулоспинальный тракт. Широкі аферентні та еферентні зв'язки структур екстрапірамідної системи між собою, двосторонні зв'язки підкіркових вузлів з корою великого мозку, особливо з її моторними областями, а також зв'язки зі структурами проміжного, середнього та довгастого мозку забезпечують широку взаємодію збуджень на нейронах, що є основою вищої інтеграції. контролю поведінкових актів
При здійсненні рухового акта частини тіла, що переміщаються, відчувають вплив інерційних сил, що порушує плавність і точність виконуваного руху. Коригують цей рух структури мозочка. У проміжну частину мозочка по колатералям кортикоспінального тракту надходить інформація про запланований рух, а також аферентація від соматосенсорної системи. В результаті формуються потоки збудження до червоного ядра та стовбурових рухових центрів, що забезпечують взаємну координацію пізніх та цілеспрямованих рухів, а також корекцію виконуваного руху. Особливо велику роль мозок грає при побудові швидких балістичних цілеспрямованих рухів (наприклад, кидання м'яча в ціль, стрибок через перешкоду, гра на фортепіано). У таких випадках корекція по ходу виконання руху неможлива через малі часові параметри, балістичний рух здійснюється тільки за заздалегідь заготовленою програмою. Вона формується в півкуль мозочка і його зубчастому ядрі на основі імпульсації, що надходить від усіх областей кори великого мозку, і фіксується в мозочку. Т.ч., протягом життя людини відбувається безперервне «навчання» мозочка зі збереженням інформації, що дозволяє пірамідній та екстрапірамідній системам сформувати необхідний комплекс рухових імпульсів, під дією яких буде виконано необхідний рух.
Структури Р. м. беруть участь у формуванні поведінки людини, яка здійснюється за принципом рефлексу, що лежить в основі всього різноманіття поведінкових актів. Аналіз закономірностей формування рефлекторних реакцій дозволив І.П. Павлову встановити фундаментальну основу навчання протягом індивідуального життя людини – умовний рефлекс. Головним становищем вчення про умовних рефлексах з'явилися уявлення про механізми замикання тимчасових зв'язків у Р. м. Результати подальших досліджень центральних механізмів формування умовного рефлексу стали основою розробки школою П.К. Анохіна системних принципів організації внутрішньомозкових процесів для формування цілеспрямованих поведінкових актів. Початковою стадією внутрішньомозкової організації поведінки будь-якого ступеня складності є аферентний синтез. Він є системним процесом зіставлення, об'єднання та відбору в структурах Г. м. різноманітних за значенням для організму численних аферентних потоків збуджень. Основними компонентами аферентного синтезу є мотиваційне збудження, механізми пам'яті, потоки обстановної та пускової аферентації. Мотиваційне збудження виникає з урахуванням внутрішньої потреби організму, наприклад харчової, питної, температурної (див. Мотивації ), у гіпоталамічних, лімбічних або ретикулярних структурах Г. м. і у вигляді висхідних активуючих впливів охоплює різні області кори великого мозку. Важливою властивістю мотиваційного порушення є його домінантність. З усього різноманіття потреб організму, що відбивають різні сторони його метаболічних змін, в даний час домінує потреба, що є найважливішою для життя індивідуума. Вона створюватиме домінуючу мотивацію, яка за фізіологічними механізмами завжди будується на основі принципу домінанти. Домінуюче збудження підвищує збудливість нейронів певних областей кори великого мозку, що призводить до збільшення їх конвергентної ємності і сприяє інтеграції інших аферентних збуджень, що приходять. Мотиваційне збудження здатне активувати вроджені механізми довгострокової пам'яті, що може реалізуватися у розгортанні жорстких, генетично визначених програм поведінки (інстинктів). Разом з тим мотиваційне збудження забезпечує фіксацію в Р. м. тих нових збуджень, що виникають при впливі на організм подразників, які сигналізують про досягнення корисних результатів поведінки. Домінуюче мотиваційне збудження легко та швидко активізує набутий індивідуальний досвід із задоволення відповідної потреби.
Сторінки: 2Розглядаючи питання класифікації умовних рефлексів, ми могли переконатися в тому, що будь-який стимул, що сприймається рецепторами та органами чуття, при підкріпленні його безумовним подразником набуває сигнальнезначення, тобто стає умовним сигналом, що викликає харчовий чи оборонний рефлекс. Павлов поставив питання: що було б з організмом, якби мало місце лише утворення умовних рефлексів, які, нашаровуючись один на одного, не давали б тварині жодної хвилини спокою? Організм виявився б буквально «пошматований» безліччю умовних рефлексів. Однак, подібного не спостерігається у житті тварин та людини. Павлов припустив, що, поряд з процесом утворення нових умовних рефлексів, йде процес гальмуваннятих старих умовних рефлексів, які не відповідають новим умовам життя, що змінилися. Так Павлов прийшов до розгляду процесів гальмуванняу вищій нервовій діяльності як процесів, протилежних до збудження. Існує безумовне (Вроджене) гальмування та умовне(Придбане, вироблене)гальмуванняумовних рефлексів. Перший - безумовне гальмування - проявляється у тварин відразу, не вимагаючи ніякого вироблення. Другий вид - умовне гальмування - передбачає шалену процедуру виробітку, тобто воно виникає за певних умов, тому і називається умовним. Неважко помітити, що гальмування, поділяючись на безумовне та умовне, ніби дзеркально повторює поділ рефлексів на безумовні та умовні. Це природно, оскільки в основу поділу рефлексів та видів гальмування покладено один принцип – їх уроджений чи набутий характер.
Розглянемо безумовне(Вроджене) гальмування. Воно підрозділяється, своєю чергою, на два різновиди: зовнішнє, чи індукційне,гальмування та позамежнегальмування. Зовнішнє гальмування виникає щоразу, коли при дії умовного сигналу раптово починає діяти інший подразник, що викликає собаку орієнтовний рефлекс «що таке?». Павлов у своїх лекціях описує, що дуже часто у собаки з виробленими міцними умовними рефлексами при демонстрації її у студентській аудиторії не вдавалося отримати слиновиділення
на умовний подразник. Аналізуючи причину невдач, Павлов дійшов висновку, що найменший рух серед студентів, нові звуки, струмінь повітря, різкий запах могли загальмувати умовний рефлекс. Це і є зовнішнєгальмування умовних рефлексів. Механізм його досить простий і пов'язаний з явищами індукції (звідси друга назва зовнішнього індукційного гальмування). При дії нових сигналів імпульси збудження від рецепторів (органів чуття) по доцентрових нервах надходять у кору великих півкуль, внаслідок чого в центрі безумовного орієнтовного рефлексу виникає вогнище збудження. Наслідком порушення цього центру є ціле віяло периферичних реакцій у вигляді повороту голови, тулуба, установки вушних раковин, фіксації поглядом нового подразника тощо. Біологічно цей рефлекс надзвичайно важливий, оскільки дозволяє оцінити ступінь небезпеки чи корисності для тваринного нового сигналу. Таким чином, координується життєдіяльність організму, у кожний момент часу здатного здійснювати лише одну діяльність, біологічно найбільш значиму.
Центр орієнтовного рефлексу в корі є домінуючим центром, збудженим набагато сильніше, ніж центр умовного рефлексу. Навколо сильнішого центру «наводиться», тобто індукується (термін запозичений з фізики); гальмування,у зону якого й попа йде центр умовного рефлексу. Внаслідок цього розвивається індукційне гальмування - тварина не реагує світ слиновиділенням (рис. 10).
46
Даний вид індукції отримав назву одночасної негативної індукції (навколо сильнішого вогнища збудження індукується процес гальмування). Одночасна негативна індукція є основою такого важливого психологічного стану, як концентрація уваги. Коли людина поглиблено займається, наприклад, розумовою роботою, то в корі великих півкуль формується стійка домінантнийвогнище збудження, що забезпечує цю діяльність, а навколо цього збудженого вогнища індукується гальмування,що робить людину нечутливою до відносно слабких різних зовнішніх впливів.
Отже, будь-який зовнішній подразник, якщо він досить сильний, може механізмом негативної індукції загальмувати умовний рефлекс. Однак, якщо новизна подразника з часом втрачається, то умовний рефлекс не гальмується цим подразником, оскільки не проявляється рефлекс «що таке?». Подібні подразники належать до так званих гаснутимгальм. Однак, є подразники, до яких тварина не може звикнути, через що вони завжди викликають індукційне гальмування і називаються постійнимигальмами. Так, вид кішки для собаки є найсильнішим подразником, який провокує мисливську поведінку, на тлі якої гальмуються умовні рефлекси на звичні команди господаря. До постійних гальм відносяться і особливі внутрішні стани тварини, фоні яких завжди гальмуються умовні рефлекси. Це, наприклад, переповнення сечового міхура, біль, сильна спрага, статеве збудження тощо.
Позамежне гальмуваннятакож належить до безумовного. Воно виникає у тварин і людини тоді, коли сила умовного подразника або частота його пред'явлення тварині занадто великі і перевершують межу працездатності крихких нервових клітин кори великих півкуль. Звідси і назва позамежнегальмування. Його можна спостерігати і за дії безумовних подразників. Якщо увімкнути водопровідний кран так, щоб капала з нього, то у собаки, що знаходиться в цій кімнаті, спочатку проявиться безумовний орієнтовний рефлекс - собака підійде до раковини, підніметься на задні лапи, зафіксує поглядом воду, що капає. Потім тварина заспокоюється, сідає поряд з раковиною і поступово звуки крапель, що падають, призводять до розвитку позамежного гальмування в тих клітинах кори великих півкуль, які були збуджені даними.
монотонним подразником. Тут ми маємо справу з послідовною негативною індукцією,оскільки осередок збудження згодом перетворюється на протилежний, гальмівний стан. Спостерігаючи за собакою, ми побачимо, що через деякий час вона згортається клубочком і засинає. У цьому випадку відбулася іррадіація гальмування з обмеженого вогнища кори великих півкуль протягом усього кору, та був і підкоркові структури. Таким чином, гальмування, як і збудження, може поширюватися по корі та підкорці, тобто іррадіювати, забезпечуючи настання сну.
Безмежне гальмування, що розвивається при дії занадто сильних або тривалих умовних і безумовних подразників, грає охоронно відновлювальнуроль, виступаючи як захід біологічного захисту, оберігаючи нервові клітини від руйнування.
Умовне гальмування
Умовне гальмуванняподіляється на 4 види: згасальне, запізнювальне, диференціювальне, умовне гальмо. Умовне гальмування, на відміну від безумовного, вимагає вироблення. Умови виробітку гальмування прямо протилежні тим, які потрібні для утворення рефлексів, тобто для того, щоб виробити умовне гальмування, потрібно скасувати підкріплення.Залежно від того, як здійснюється непідкріпленняумовного сигналу і розрізняють перелічені вище види умовного гальмування.
Згасальне гальмування.Якщо у собаки вироблений умовний слиновидільний рефлекс, його можна згасити, скасувавши звичне харчове підкріплення. Наведемо приклад дослідів Павлова із погашенням харчового умовного рефлексу, виробленого на звуки метронома.

48
З цього протоколу досвіду видно, що скасування звичного підкріплення призводить до досить швидкого гасіння умовного рефлексу, проте до кінця "від рефлекс, як правило, не згасає. Далі, у голодного собаки харчовий умовний рефлекс згасити важче, ніж у ситої Ці особливості процесу гасіння легко зрозумілі.
Що стоїть за гасінням умовного рефлексу? Руйнуєтьсятимчасовий зв'язок або тільки гальмується?Важливою властивістю всіх умовних рефлексів є їхня здатність до мимовільному відновленню.Якщо собаці з згашеним умовним рефлексом наступного дня запропонувати як умовний подразник удари метронома, то у тварини відновиться умовне слиновиділення. Це доводить, що при погашенні умовного рефлексу у попередній день він не руйнується. Іншим доказом гальмування, але з руйнації тимчасових зв'язків при процедурі гасіння шляется використання раптового сильного подразника і натомість згаслого умовного рефлексу. Цей раптовий новий подразник ніби розгальмує згасальний процес, тобто, за словами Павлова, викличе гальмування гальмування,внаслідок чого на умовний сигнал знову буде слиновиділення. Якщо у собаки створити сильну харчову мотивацію, тобто не годувати її деякий час, то умовний сигнал, на який умовну реакцію було погашено, знову стає ефективним. Таким чином, при погашенні тимчасові зв'язки в корі великих півкуль не руйнуються, а лише гальмуються.
Запізнювальне гальмування.Цей вид гальмування проявляється при виробленні рефлексів, що запізнюються, де умовний подразник не відразу підкріплюється безумовним, а через 1-2 хв після початку його дії і умовного сигналу. У цьому рефлексі Павлов розрізняв дві фази -не діяльнуі діяльну.Перша недіяльна фаза характеризується відсутністю умовної реакції протягом 1-2 хв після початку дії умовного сигналу. У другій, діяльній фазі, спостерігається виділення слини. Аналізуючи недіяльну фазу, Павлов дійшов висновку, що у її основі лежить гальмування, назване їм запізнюється.Доказом цього є процес розгальмовування за допомогою нових сторонніх подразників. Якщо на етапі недіяльної фази увімкнути новий сигнал,
викликає у собаки рефлекс «що таке?», то спостерігається описане вище гальмування гальмування,тобто розгальмовування, внаслідок чого у тварин починає виділятися слина. Цей досвід показує, що відсутність реакції в недіяльній фазі рефлексу, що запізнюється, відображає наявність активного процесу гальмування. Можна розгальмувати недіяльну фазу, створивши сильну харчову мотивацію тварини. У разі, якщо собака голодна, то слиновиділення починається відразу ж за умови пред'явлення умовного сигналу. Запізнювальне гальмування відіграє важливу роль у життєдіяльності тварин, що можна бачити на прикладі харчодобувної поведінки хижаків. Вистежуючи видобуток протягом кількох годин, хижак здійснює масу рухових рефлексів (умовних та безумовних), тобто здійснюється руховий компонент харчової поведінки. Разом з тим, вегетативний компонент у формі виділення слини, шлункового соку загальмований, і лише коли видобуток наздогнутий, починаються процеси умовно- та безумовнорефлекторного виділення травних соків, що забезпечують хімічну переробку їжі. Відставленість цих процесів біологічно виправдана, оскільки передчасне вироблення наприклад, шлункового соку, що містить соляну кислоту, призвела б до утворення виразок у шлунково-кишковому тракті.
Диференціювальне гальмування.Цей вид гальмування лежить в основі розрізнення близьких подразників. Якщо, наприклад, у собаки вироблений умовний харчовий рефлекс на ноту до третьої октави, то при пред'явленні тварині будь-якої іншої ноти у неї спочатку виявлятиметься ця ж умовна реакція. Однак, потім, оскільки їжею підкріплюється тільки нота ДО, всі інші звуки перестануть викликати слиновиділення. диференціювальногогальмування. Довести, що відсутність реакції пов'язане саме з розвитком гальмування, можна застосувати прийом розгальмовування за допомогою сторонніх сигналів, або створивши сильну харчову мотивацію у собаки. При виробленні диференціювання необхідно починати з подразників, які сильно відрізняються один від одного. Так, у собаки умовним подразником було зроблено коло, яке підкріплювали їжею. Це коло стали чергувати з еліпсом із співвідношенням осей 8:9, тобто за формою дуже близьким до кола. Еліпс їжею не підкріплювали. Спроби добитися розрізнення
цих двох дуже схожих постатей не призвели до позитивних результатів. У собаки стався зрив вищої нервової діяльності, що виявилося в агресії – тварина почала зривати з себе приладчики, рватися зі верстата, гарчати на експериментатора тощо. Отже, це завдання на розрізнення було для собаки непосильним. Разом про те, можна домогтися вироблення диференціювального гальмування даний еліпс за іншого підході. Спочатку собаці пред'являють коло, що підкріплюється, їжею та еліпс із співвідношенням осей 4:8 (сильно відрізняється від кола), що не підкріплюється їжею. Диференціювання виробляється через кілька поєднань – тварина позитивно реагує на коло та не реагує на еліпс. Потім береться це коло і еліпс із співвідношенням осей 5:8, і процедура повторюється. Потім до кола додається еліпс із співвідношенням осей 6:8. Після швидкого вироблення диференціювання цей еліпс переходять, нарешті, до протиставлення кола і еліпса із співвідношенням осей 7:8. Використовуючи метод поступовоївироблення диференціювального гальмування, всього після 18 поєднань кола, що підкріплюється їжею, і еліпса, що не підкріплюється, вдалося домогтися розрізнення тваринам кола і еліпса з співвідношенням осей 7:8.
Диференціювальне гальмування, забезпечуючи тонке розрізнення тваринами та людиною близькоспоріднених подразників, сприяє пеціалізаціїумовних рефлексів, тобто точного і правильного реагування зовнішні стимули.
Умовне гальмо.Цей вид гальмування може спостерігатися за умов умовних рефлексів II порядку. Згадаймо процедуру вироблення цих рефлексів. Спочатку виробляється умовний рефлекс I порядку, наприклад, світ, що підкріплюється їжею. В результаті декількох поєднань світло набуває сигнального значення, тобто спалах лампочки супроводжується умовним слиновиділенням. Потім собаці пропонують новий (подразник, наприклад, удари метронома, і підкріплюють їх світлом, внаслідок чого метроном також набуває сигнального значення, тобто викликає слиновиділення, хоча й слабше, ніж світло. Виявилося, що за такої комбінації новогоі звичного умовнихсигналів який завжди виробляється умовний рефлекс II порядку. Якщо новий подразник дуже сильний або проміжок часу між новим та звичним подразником занадто малий, то виробляється умовне гальмона цю комбінацію, тобто поєднання метронома зі світлом або один метроном не будуть
супроводжуватися слиновиділенням. Причиною вироблення умовного гальма замість умовного рефлексу II порядку є надмірна сила нового подразника чи занадто маленька пауза між подразниками. У цих випадках собака сприймає комбінацію як новий непідкріплюваний їжею комплекс,оскільки звичний подразник (світло) маскується новим сильнішим подразником.
Механізм умовного гальмування.Питання, які процеси забезпечують гальмування умовних рефлексів, Павлов називав «проклятим» питанням, оскільки гальмування, на відміну збудження, зовні не виявляє і тому важко піддається вивченню. Розглянемо сучасні гіпотези про механізм умовного гальмування. Перша з них пов'язана з дослідженнями Асратяна Е.А.і допомагає відповісти на запитання, де,у яких структурах умовного рефлексу виникають гальмівні процеси. У собаки виробляли різні умовні рефлекси (харчові та оборонні) на умовні сигнали. При цьому експерименти проводили у двох різних кімнатах. Виявилося, що на той самий подразник, наприклад, світло, можна, підкріплюючи його в першоюкімнаті їжею, виробити харчовий умовний рефлекс, а в іншій кімнаті, підкріплюючи світло роздратуванням лапи струмом, умовний оборонний рефлекс. Собака, перебуваючи в першій кімнаті, реагуватиме на світло слиновиділенням, перебуваючи в другій кімнаті оборонною руховою реакцією. Ці досліди отримали назву дослідів з перемиканням умовних рефлексів.Вони демонструють роль
 подразників обстановкиконкретних кімнат, які самі собою не викликають умовних рефлексів, але відіграють роль перемикачів,тобто готують тварину до конкретноїумовної реакції на той самий подразник. У цього ж собаки у першій кімнаті вироблявся умовний харчовий рефлекс на удари метронома. Потім у тварини проводили погашення умовного слиновидільного рефлексу на світ у першій кімнаті. Відміна
подразників обстановкиконкретних кімнат, які самі собою не викликають умовних рефлексів, але відіграють роль перемикачів,тобто готують тварину до конкретноїумовної реакції на той самий подразник. У цього ж собаки у першій кімнаті вироблявся умовний харчовий рефлекс на удари метронома. Потім у тварини проводили погашення умовного слиновидільного рефлексу на світ у першій кімнаті. Відміна
підкріплення світла їжею призвела до розвитку згасателъного гальмування.Виникає питання, де воно локалізовано?На рис. 11 представлена схема умовного рефлексу, що включає центр умовного подразника, центр безумовного підкріплення та тимчасовий зв'язок між цими центрами.
Таким чином, при погашенні умовного рефлексу гальмування може локалізуватися або в центрі умовного подразника (1), або в центрі безумовного підкріплення (3), або в часі (2). Для з'ясування питання Асратян використовував вироблені у собаки різноманітні умовні рефлекси. Так, у першій кімнаті після того, як собака перестав реагувати на світ слиновиділенням, включили метроном, на який раніше у цього собаки був також вироблений умовний слиновидільний рефлекс. Виявилося, що тварина реагує на метроном слиновиділенням. Отже, при процедурі погашення умовного рефлексу світ центр безумовного підкріплення не виявився загальмованим. Далі собаку вводили в другу кімнату і знову включали світло, на яке в цій кімнаті собака раніше виробив умовний оборонний рефлекс. Виявилося, що цей умовний рефлекс на світло добре проявляється, що дозволяє зробити висновок про те, що при погашенні умовного харчового рефлексу на світ у першій кімнаті гальмування не локалізується в центрі умовного сигналу. Таким чином, погашення умовного рефлексу не призводить до розвитку гальмування ні у центрі умовного подразника, ні у центрі безумовного підкріплення. Отже, місцем, де спочатку локалізується гальмування, є сама тимчасовий зв'язок.Такий висновок був зроблений Асратяном виходячи з вищеописаних експериментів. Якщо після погашення умовного рефлексу продовжувати пред'являти собаці світло, не підкріплюючи його їжею, то тварина з часом перестає реагувати і на метроном, і навіть на їжу, що свідчить про іррадіаціїгальмування із системи нейронів, що утворюють тимчасовий зв'язок, на центри умовного та безумовного подразника. Більш того, тварина може заснути при пред'явленні світлових сигналів, що доводить, що доводить іррадіацію гальмування у всій корі і підкіркових структур.
Таким чином, спочатку гальмування розвивається в системі нейронів, що утворюють тимчасовий зв'язок, а потім може іррадіювати, охоплюючи всю структуру умовного рефлексу, а також всю кору та підкірку, що призводить до сну.
Своєрідні погляди природу умовного гальмування належать П.К. Анохіна. Спостерігаючи за поведінкою собаки під час погашення харчових умовних рефлексів, Анохін звернув увагу на те. що скасування звичного підкріплення супроводжується різними руховими реакціями тварини у формі повороту голови у різні боки, принюхування, переступання з лапи на лапу тощо. Анохін назвав цей стан собаки важким станомабо біологічно негативною реакцією,центр якої у корі великих півкуль сильно збуджений і забезпечує описані вище рухові реакції (рис. 11 Б). За поясненням Анохіна, тварина як би «неприємно здивована» тим, що за умовним сигналом не слідує позитивного підкріплення. Центр біологічно негативної реакції, що виник у корі, за законами індукціїгальмує умовний рефлекс, чим пояснюється механізм згасального гальмування. Таким чином, згідно з поглядами Анохіна, умовне гальмування розвивається за тими ж механізмами. одночасної негативної індукції,що і зовнішнєгальмування, описане вище.
Теорія Анохіна про індукційний характер умовного гальмування переконлива при поясненні гальмування харчових,але з оборонних умовних рефлексів. Справді, важко, користуючись термінологією Анохіна, пояснити механізм погашення умовного оборонного рефлексу, наприклад, світ. Скасування больового підкріплення призводить до того, що тварина перестає реагувати світ відсмикуванням тієї лапи, яка дратувалася електричним струмом. Чи можна пояснити це гальмування виникненням біологічно негативної реакції чи тяжкого стану? Навряд чи тварина неприємно здивована тим, що світло не підкріплюється больовим роздратуванням.
П.С. Купалов запропонував універсальнішу схему гальмування, що пояснює гальмування будь-яких видів рефлексів. Суть його концепції в тому, що при скасуванні підкріплення, незалежно від того, харчове воно чи больове, у тварини виникає орієнтовний рефлекс «що таке?», центр якого за законами негативноюіндукції гальмує центр умовного рефлексу
Таким чином, запропонована Анохіним та Купаловим концепція пояснює механізм зовнішньогоі внутрішнього гальмуванняіз загальних позицій -розвитку негативної індукціїчерез виникнення нових вогнищ збудження в корі великих півкуль.
Слід зазначити, що проблема умовного гальмування неспроможна вважатися повністю вирішеною, зокрема, незрозумілими залишаються нейрофізіологічні основи гальмування. Успіхом слід вважати виявлення гальмівного медіатора - гамма-аміномасляної кислоти, виділення якої пресинаптичними мембранами блокує проведення збудження в центральній нервовій системі.
є потужне вогнище збудження в корі головного мозку, що викликає гальмування в ділянках кори, що оточують його, за законом негативної індукції.
Зовсім інший вид неуважності спостерігається в тих випадках, коли людина не в змозі ні на чому довго зосередитися, коли вона постійно переходить від одного об'єкта чи явища до іншого, ні на чому не затримуючись. Цей вид розсіяності називається справжньою неуважністю.Довільна увага людини, яка страждає на справжню розсіяність, відрізняється крайньою нестійкістю і відволіканням. Фізіологічно справжня розсіяність пояснюється недостатньою силою внутрішнього гальмування. Порушення, що виникає під впливом зовнішніх сигналів, легко поширюється, але важко концентрується. В результаті в корі мозку розсіяної людини створюються нестійкі осередки збудження.
Причини справжньої розсіяності різноманітні. Ними можуть бути загальний розлад нервової системи, захворювання крові, нестача кисню, фізична чи розумова втома, тяжкі емоційні переживання. Крім того, однією з причин справжньої розсіяності може бути значна кількість отриманих вражень, а також невпорядкованість захоплень та інтересів.
14.4. Розвиток уваги
Увага, як більшість психічних процесів, має свої етапи розвитку. У перші місяці життя у дитини відзначається наявність лише мимовільної уваги. Дитина спочатку реагує лише зовнішні подразники. Причому це відбувається тільки у разі їх різкої зміни, наприклад, при переході з темряви до яскравого світла, при раптових гучних звуках, при зміні температури тощо.
Починаючи з третього місяця дитина все більше цікавиться об'єктами, тісно пов'язаними з його життям, тобто найближчими до нього. У п'ять-сім місяців дитина вже може досить довго розглядати якийсь предмет, обмацувати його, брати в рот.
Особливо помітним є прояв його інтересу до яскравих та блискучих предметів. Це дозволяє говорити про те, що його мимовільна увага вже досить розвинена.Зачатки довільної уваги зазвичай починають виявлятися до кінця першого – початку другого року життя. Можна припустити, що виникнення та формування довільної уваги пов'язане із процесом виховання дитини. Люди, що оточують дитину, поступово привчають її виконувати не те, що їй хочеться, а те, що їй потрібно робити. На думку М. Ф. Добриніна, в результаті виховання діти змушені звертати увагу на необхідну від них дію, і поступово у них, поки що у примітивній формі, починає виявлятися свідомість.
Велике значення у розвиток довільного уваги має гра. У процесі гри дитина вчиться координувати свої рухи згідно з завданнями; ри та спрямовувати свої дії відповідно до її правил. Паралельно
Глава 14. Увага 371
з довільною увагою на основі чуттєвого досвіду розвивається і мимовільна увага. Знайомство з дедалі більшою кількістю предметів і явищ, поступове формування вміння розбиратися у найпростіших стосунках, постійні розмови з батьками, прогулянки з ними, ігри, в яких діти наслідують дорослих, маніпулювання іграшками та іншими предметами – все це збагачує досвід дитини, а разом з тим розвиває його інтереси та увагу.
Основною особливістю дошкільника є те, що його довільна увага є досить нестійкою. Дитина легко відволікається на сторонні подразники. Його увага надмірно емоційна, - він ще погано володіє своїми почуттями. При цьому мимовільна увага досить стійка, тривала і зосереджена. Поступово шляхом вправ та вольових зусиль у дитини формується здатність керувати своєю увагою.
Особливого значення у розвиток довільного уваги має школа. У процесі шкільних занять дитина привчається до дисципліни.
У нього формується посидючість, здатність контролювати свою поведінку. Слід зазначити, що у шкільному віці розвиток довільної уваги проходить певні стадії. У перших класах дитина не може повністю контролювати свою поведінку на уроках. У нього, як і раніше, переважає мимовільна увага. Тому досвідчені вчителі прагнуть зробити свої заняття яскравими, захоплюючими увагу дитини, що досягається періодичною зміною форми подачі навчального матеріалу. При цьому слід пам'ятати, що у дитини у цьому віці мислення здебільшого наочно-подібне. Тому, щоб привернути увагу дитини, виклад навчального матеріалу має бути гранично наочним.У старших класах довільна увага дитини досягає вищого рівня розвитку. Школяр вже може досить тривалий час займатися певним видом діяльності, контролювати свою поведінку. Проте слід пам'ятати, що у якість уваги впливають як умови виховання, а й особливості віку. Так, фізіологічні зміни, що спостерігаються у віці 13-15 років, супроводжуються підвищеною стомлюваністю та дратівливістю та в деяких випадках призводять до зниження характеристик уваги. Це зумовлено як фізіологічними змінами організму дитини, а й значним зростанням потоку сприймається інформації та вражень школяра.
Л. С. Виготський намагався у межах своєї культурно-історичної концепції простежити закономірності вікового розвитку уваги. Він писав, що з перших днів життя дитини розвиток його уваги відбувається в середовищі, що включає так званий подвійний ряд стимулів,викликають увагу. Перший ряд - це навколишні дитини предмети, які своїми яскравими, незвичайними властивостями приковують його увагу. З іншого боку - це мова дорослої людини, слова, що вимовляються їм, які спочатку виступають у вигляді стимулів-вказівок, що спрямовують мимовільну увагу дитини. Довільна увага виникає з того, що люди, що оточують дитину, починають за допомогою низки стимулів і засобів спрямовувати увагу дитини, керувати її увагою, підкоряти її своїй волі і тим самим дають до рук дитини ті засоби, за допомогою
Осередок застійного збудження, що має певний іпохондричний зміст, може утворитися і за надмірної фіксації уваги на діяльності будь-якого органу. Тим самим завдяки наявності зворотного зв'язку між мозком і будь-яким внутрішнім органом, будь-якою ділянкою нашого тіла не тільки зростає сила відчуття, але здатна змінюватися і функція даного утворення (згадаймо про вищезгадані вісцеральні умовні рефлекси).
Слід також нагадати, що при невротичних станах нервова система стає більш менш астенізованою, що робить її і більш чутливою. Звідси можливість усвідомлення подразнень, що надходять із внутрішнього середовища організму в центральну нервову систему, які в нормі не відчуваються: перистальтика шлунка, кишечника, відчуття биття серця в спокійному стані та ін.
При ипохондрическом неврозі ці явища різко посилюються внаслідок вираженої зосередженості уваги хворого цих відчуттях.
Як вказують Н. Ф. Суворов, В. Б. Захаржевський та ін (1980), при неврозі первинно порушується функція кори головного мозку. При цьому підкіркові вегетативні центри виходять з-під контролю, що викликає розлад діяльності внутрішніх органів. З іншого боку, відновлення стану кори призводить до нормалізації порушених механізмів.
Однак при дії психотравмуючих обставин можуть розвиватися розлади різних систем організму без наявності неврозу. Це звані психосоматичні захворювання.
У цих випадках надмірна напруга нервової системи здатна викликати гіпертонічну хворобу, виразку шлунка або дванадцятипалої кишки, діабет та ін., як видно, при вже наявній схильності до цих захворювань. Велику роль при цьому може відігравати сильне збудження судинного центру, схильність до спастичного стану кровоносних судин, наприклад певних ділянок травного тракту та ін.
Звичайно, не виключено поєднання неврозу та психосоматичного захворювання. У подібних випадках, крім суворого спостереження за виконанням хворим усіх лікувальних призначень, основна увага середнього медперсоналу зосереджується на роз'ясненні та навіювання хворому, що під дією лікування його стан обов'язково поліпшуватиметься.
Хворого потрібно переконувати в тому, що ніякого істотного органічного захворювання у нього немає, що про це свідчать всі дослідження, що проводилися. Його хворобливий стан - це результат колишнього в минулому захворювання, або нервово-психічних перенапруг, або будь-яких інших шкідливостей, що послабили його організм, а на цьому тлі у нього розвинулися хворобливі уявлення про наявність тяжкого соматичного захворювання.
Слід врахувати, що в процесі лікування можуть бути тимчасові погіршення у зв'язку, наприклад, зі зміною барометричного тиску, порушенням нічного сну, будь-яким додатковим переживанням та ін. Іноді звертають увагу хворого на те, що він став виглядати краще, молодшим і бадьорішим.
«Догляд за хворими з невротичними станами»,
Л.Г. Перший

У 2018 році Різдвяний пост розпочнеться 28 листопада. У цей період православні віруючі готуються до зустрічі Різдва.

Створення сім'ї – мрія більшості жінок. Вони хочуть мати люблячого чоловіка і купу дітей. Тільки от не завжди стосунки...

Ця стаття містить: найсильніша молитва від розлучення - інформація взята зі всіх куточків світла, електронної мережі та...

Інформаційний сайт про ікони, молитви, православні традиції. Молитва від скандалів та сварок у сім'ї, з чоловіком, з дітьми...

Який же Новий рік без шампанського, мандарин, «Олів'є», заливного та всіма улюбленої «Оселедця під шубою». Ось із останнім...

Підготуємо необхідні інгредієнти для печива. Перше, що потрібно зробити - це поставити воду кип'ятитися. Нам необхідний...

Чи можна оформити працівника посаду фінансовий директор - головний бухгалтер. Головний бухгалтер стверджує, що...

Керівник невеликого бізнесу може вести бюджет самостійно. ПЕРЕВІРЕНО! Якщо займатися веденням...

Створення нових проектів передбачає попереднє економічне обґрунтування їхньої доцільності, наступне...
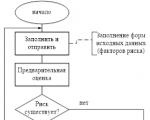
Звітність формується РМ, проходить узгодження (схвалення) Комітету з ризиків за Правління та передається на...

На краю великого луга, на довгій смарагдовій травинці жила крихітна Божа Корівка. Маленька Божа...

Нині досить поширене звернення людини до зірок. За допомогою гороскопу людина може дізнатися...

Діловий чи приятельський. Якщо ж переслідували незнайомку - значить рівень вашої довіри до жіночої статі.

по соннику ФрейдаЯкщо вам снилося, як ви ловили рибу, значить, у реальному житті ви насилу можете відключитися...

Створення сім'ї – мрія більшості жінок. Вони хочуть мати люблячого чоловіка і купу дітей. Тільки от не завжди...

Ця стаття містить: найсильніша молитва від розлучення - інформація взята зі всіх куточків світла, електронної...