Готвене на малки октоподи: тайните на най-добрите готвачи
Римляните и древните гърци много обичали октоподите и постоянно ги ловували. Любовта е минала през вековете и сега...
Зад и над носа е ниша на прозореца на вестибюла (fenestra vestibuli),с форма на овал, удължен в предно-задна посока, с размери 3 на 1,5 mm. Прозорецът на вестибюла е затворен основата на стремето (basis stapedis),прикрепен към краищата на прозореца
Ориз. 5.7.Медиалната стена на тъпанчевата кухина и слуховата тръба: 1 - нос; 2 - стреме в нишата на прозореца на вестибюла; 3 - кохлеарен прозорец; 4 - първо коляно на лицевия нерв; 5 - ампула на страничния (хоризонтален) полукръгъл канал; 6 - барабанна струна; 7 - стапедиален нерв; 8 - югуларна вена; 9 - вътрешна каротидна артерия; 10 - слухова тръба
като се използва пръстеновиден лигамент(lig. annulare stapedis).В областта на задно-долния ръб на носа има прозоречна ниша за охлюви (fenestra Cochleae),продължителен вторично тъпанче (membrana tympani secundaria).Нишата на прозореца на кохлеята е обърната към задната стена на тъпанчевата кухина и е частично покрита от проекцията на задно-долния наклон на промонториума.
Непосредствено над прозореца на вестибюла в костния фалопиев канал преминава хоризонталното коляно на лицевия нерв, а отгоре и отзад има издатина на ампулата на хоризонталния полукръгъл канал.
Топография лицев нерв (n. facialis, VII черепномозъчен нерв)има важно практическо значение. Присъединяване към н. statoacousticusИ н. intermediusвъв вътрешния слухов канал, лицевият нерв преминава по дъното му, в лабиринта се намира между вестибюла и кохлеята. В лабиринтния участък той се отклонява от секреторната част на лицевия нерв голям каменист нерв (n. petrosus major),инервиращ слъзна жлеза, както и лигавичните жлези на носната кухина. Преди да излезете в тъпанчевата кухина, над горния ръб на прозореца на вестибюла има коленчат ганглий (ganglion geniculi),при които се прекъсват вкусовите сетивни влакна на междинния нерв. Преходът на лабиринтната секция към тимпаничната секция се обозначава като първи род на лицевия нерв.Лицевият нерв, достигащ до издатината на хоризонталния полукръгъл канал на вътрешната стена, на ниво пирамидална височина (eminentia pyramidalis)променя посоката си на вертикална (второ коляно)преминава през стиломастоидния канал и през едноименния форамен (за. stylomastoideum)се простира до основата на черепа. В непосредствена близост до пирамидалното възвишение лицевият нерв отделя клон към стапедиален мускул (m. stapedius),тук се отклонява от ствола на лицевия нерв барабанна струна (chorda tympani).Той преминава между чука и инкуса през цялата тъпанчева кухина от над тъпанчето и излиза през fissura petrotympanica (s. Glaseri),придавайки вкусови влакна на предните 2/3 на езика отстрани, секреторни влакна на слюнчена жлезаи влакна към нервните съдови плексуси. Стената на канала на лицевия нерв в тъпанчевата кухина е много тънка и често има дехисценция, което определя възможността за разпространение на възпалението от средното ухо към нерва и развитието на пареза или дори парализа на лицевия нерв. Различни локализации на лицевия нерв в тъпанчевата и мастоидната
Средното ухо, auris media, се състои от тъпанчевата кухина и слухова тръбасвързващ тъпанчевата кухина с назофаринкса.
Тимпаничната кухина, cavitas tympanica, се намира в основата на пирамидата темпорална костмежду външния слухов канал и лабиринта (вътрешното ухо). Съдържа верига от три малки кости, които предават звукови вибрации от тъпанчето към лабиринта.
Тъпанчевата кухина е с много малък размер (обем около 1 кубичен см) и прилича на тамбура, поставена на ръба й, силно наклонена към външната страна. Ушния канал. В тъпанчевата кухина има шест стени:
1. Страничната стена на тимпаничната кухина, paries membranaceus, се образува от тимпаничната мембрана и костната плоча на външния слухов канал. Горната куполообразна разширена част на тимпаничната кухина, recessus membranae tympani superior, съдържа две слухови осикули; главата на чука и инкуса. В случай на заболяване патологични промениСредното ухо е най-голямо в този рецесус.
2. Медиалната стена на тимпаничната кухина е в съседство с лабиринта и затова се нарича лабиринтна, paries labyrinthicus. Има два прозореца: кръгъл прозорец, прозорец на кохлеята - fenestra cochleae, водещ в кохлеята и покрит от membrana tympani secundaria, и овален прозорец, прозорец на преддверието - fenestra vestibuli, отварящ се в vestibulum labyrinthi. Основата на третата слухова костица, стремето, се вкарва в последния отвор.
3. Задната стена на тъпанчевата кухина, paries mastoideus, носи издигане, eminentia pyramidalis, за настаняване на m.stapedius. Recessus membranae tympani superior продължава отзад в пещерата на мастоидния процес, antrium mastoideum, където се отварят въздушните клетки на последния, cellulae mastoideae. Antrum mastoideum е малка кухина, изпъкнала към мастоидния израстък, от чиято външна повърхност е отделена от слой кост, граничещ със задната стена на слуховия канал непосредствено зад spina suprameatica, където пещерата обикновено се отваря по време на нагнояване в мастоидния процес.
4. Предната стена на тъпанчевата кухина се нарича paries caroticus, тъй като вътрешната каротидна артерия е близо до нея. В горната част на тази стена има вътрешен отвор на слуховата тръба, ostium tympanicum tubae auditivae, който се отваря широко при новородени и малки деца, което обяснява честото проникване на инфекция от назофаринкса в кухината на средното ухо и по-нататък в череп.
5. Горна стенаТъпанчевата кухина, paries tegmentalis, съответства на предната повърхност на пирамидата на тимпаничната кухина и разделя тъпанчевата кухина от черепната кухина.
6. Долната стена или дъното на тъпанчевата кухина, paries jugularis, е обърната към основата на черепа в съседство с fossa jugularis.
Три малки слухови костици, разположени в тъпанчевата кухина, се наричат малеус, инкус и стреме.
1. Malleus, malleus, е снабден със заоблена глава, caput mallei, която през шията, collum mallei, е свързана с дръжката, manubrium mallei.
2. Инкусът, incus, има тяло, corpus incudis, и два отклоняващи се процеса, от които един по-къс, crus breve, е насочен назад и лежи върху ямката, а другият дълъг процес, crus longum, върви успоредно на дръжката на чукчето медиално и отзад от него и в края му има малко овално удебеляване, processus lenticularis, съчленено със стремето.
3. Стремето, стремето, по своята форма оправдава името си и се състои от малка глава, caput stapedis, носеща ставната повърхност на incus processus lenticularis и два крака: предната, по-права, crus anterius и задната, по-извит, crus posterius, които са свързани с овална плоча, basis stapedis, вмъкната в прозореца на вестибюла.
На ставите слухови костиципомежду си се образуват две истински стави с ограничена подвижност: articulatio incudomallearis и articulatio incudostapedia. Плочата на стремето е свързана с ръбовете на fenestra vestibuli чрез съединителна тъкан, синдесмоза тимпаностапедия
Слуховите костици също са подсилени от няколко отделни връзки. Като цяло и трите слухови костици представляват повече или по-малко подвижна верига, преминаваща през тъпанчевата кухина от тъпанчето до лабиринта. Подвижността на осикулите постепенно намалява в посока от чукчето към стремето, което предпазва спиралния орган, разположен в вътрешно ухо, от прекомерни удари и остри звуци.
Веригата от осикули изпълнява две функции: 1) костна проводимост на звука и 2) механично предаване звукови вибрацииДа се овален прозорецвестибюл, fenestra vestibuli.
Последната функция се осъществява благодарение на два малки мускула, свързани със слуховите осикули и разположени в тъпанчевата кухина, които регулират движенията на веригата от осикули. Един от тях, m.tensor tympani, е вграден в semicanalis m.tensoris tympani, компонент горна част canalis musculotubarius на темпоралната кост; сухожилието му е прикрепено към дръжката на чука близо до шията. Този мускул, дърпайки назад дръжката на чука, напряга тъпанчето. В този случай цялата система от осикули се измества навътре и стремето се притиска към вестибюла на fenestra. Мускулът се инервира от третия клон тригеминален нервчрез клон на n.tensoris tympani. Друг мускул, m.stapedius, е поставен в eminentia pyramidalis и е прикрепен към задния крак на стремето в главата. Функционално този мускул е антагонист на предишния и произвежда обратно движение на осикулите в средното ухо, в посока от прозореца на вестибюла. Мускулът получава своята инервация от n.facialis, който, минавайки до него, води до малък клон, n.stapedius.
Като цяло функцията на мускулите на средното ухо е разнообразна:
1) поддържане на нормалния тонус на тъпанчето и веригата от слухови осикули;
2) защита на вътрешното ухо от прекомерна звукова стимулация и
3) приспособяване на звукопроводящия апарат към звуци различни силни странии височини. Основният принцип на средното ухо като цяло е звукопроводимостта от тъпанчето до овалния прозорец на вестибюла, fenestra vestibuli.
Слуховата или евстахиевата тръба, tuba auditiva (Eustachii; оттук и името на възпалението на тръбата - евстахит), служи за достъп на въздух от фаринкса в тъпанчевата кухина, което поддържа баланс между налягането в тази кухина и външното атмосферно налягане, което е необходимо за правилното преминаване към лабиринтните вибрации на тъпанчето.
Слуховата тръба се състои от костни и хрущялни части, които са свързани помежду си. На мястото на тяхното свързване (isthmus tubae) тръбният канал е най-тесен. Костната част на тръбата, започваща в тъпанчевата кухина с отвор, ostium tympanicum tubae auditivae, заема долната по-голяма част на мускулно-тръбния канал (semicanalis tubae auditivae) на темпоралната кост. Хрущялната част, която е продължение на костната, е изградена от еластичен хрущял.
Надолу тръбата завършва на страничната стена на назофаринкса с фарингеалния отвор, ostium pharyngeum tubae auditivae, а ръбът на хрущяла, изпъкнал във фаринкса, образува torus tubarius. Лигавицата, покриваща слуховата тръба, е покрита с ресничест епител и съдържа лигавични жлези, glandulae tubariae и лимфни фоликули, които се натрупват в голям брой в устието на фаринкса (тубарна тонзила). Влакната на m.tensor veli palatini произхождат от хрущялната част на тръбата, в резултат на което, когато този мускул се свива по време на преглъщане, луменът на тръбата може да се разшири, което улеснява навлизането на въздух в тъпанчевата кухина.
Съдове и нерви на средното ухо. Артериите произлизат главно от a.carotis externa. Множество съдове проникват в тъпанчевата кухина от нейните клонове: от a.auricularis posterior, a.maxillaris, a pharyngea ascendens, както и от ствола на a.carotis interna, когато преминават през неговия канал. Вените придружават артериите и се вливат в plexus pharyngeus, vv.meningeae mediae и v.auricularis profunda. Лимфни съдовеСредното ухо отива отчасти до възлите на страничната стена на фаринкса, отчасти до лимфните възли зад ушната мида.
Нерви: лигавицата на тъпанчевата кухина и слуховата тръба се захранва от сетивни клонове от n.tympanicus, който се простира от ganglion inferius на глософарингеалния нерв. Заедно с клоните на симпатиковия плексус на вътрешната артерия те образуват тимпаничния сплит, plexus tympanicus. Горното му продължение е n.petrosus minor, отиващ към ganglion oticum. Двигателните нерви на малките мускули на тъпанчевата кухина са посочени в тяхното описание.
4.52. Вътрешно ухо: общ план на структурата, диаграма на звуковата вълна.
Вътрешното ухо или лабиринтът се намира в дебелината на пирамидата на темпоралната кост между тъпанчевата кухина и вътрешния слухов канал, през който n. излиза от лабиринта. vestibulocochlearis. Разграничете костни и мембранни лабиринти,а последният лежи вътре в първия.
Костният лабиринт, labyrinthus osseus, е поредица от малки взаимосвързани кухини, стените на които се състоят от компактна кост. Той разграничава три секции: преддверие, полукръгли канали и кохлея; кохлеята лежи отпред, медиално и малко по-ниско от вестибюла, а полукръглите канали лежат отзад, латерално и отгоре на него.
1. вестибюл, вестибулумаобразувайки средната част на лабиринта, представлява малка кухина с приблизително овална форма, свързана отзад с пет отвора с полукръглите канали, а отпред с по-широк отвор с кохлеарния канал. На страничната стена на вестибюла, обърната към тъпанчевата кухина, има отвор, който вече ни е известен, fenestra vestibuli, зает от плочата на стремето. Друг отвор, fenestra cochleae, покрит с membrana tympani secundaria, се намира в началото на кохлеята. С помощта на гребен, Криста вестибули,преминавайки по вътрешната повърхност на средната стена на вестибюла, кухината на последния е разделена на две вдлъбнатини, от които задната, свързваща се с полукръглите канали, се нарича рецесус елиптик,и предната, най-близо до кохлеята, - recessus sphericus.В recessus ellipticus произхожда от малък отвор, apertura interna aqueductus vestibuli, акведуктът на вестибюла, преминаващ през костното вещество на пирамидата и завършващ на задната му повърхност. Под задния край на мидата на долната стена на преддверието има малка ямка, рецесус кохлеарис,съответстващ на началото на мембранозния ход на кохлеята.
2. Костни полукръгли канали, сanales miles semicirculares ossei,-
три сводести костни прохода, разположени в три взаимно перпендикулярни равнини. Преден полукръгъл канал,
canalis semicircularis anterior,разположени вертикално под прав ъгъл спрямо
ос на пирамидата на темпоралната кост, заден полукръгъл канал, canalis semicir
cularis posteriorсъщо вертикални, разположени почти успоредно на гърба
повърхността на пирамидата и страничния канал, canalis semicmrculares lateralis,
лежи хоризонтално, изпъкнал към тъпанчевата кухина. Всеки го има
каналът има два крака, които обаче се отварят само в вестибюла
пет дупки, тъй като съседните краища на предния и задния канал
са свързани в един общ крак, крус комуна.По един от всеки крак
каналът преди вливането му в преддверието образува разширение, т.нар
произведени от ампулата. Кракът с ампулата се нарича crus ampullare,и крака без
разширения - crus simplex.
3. Охлюв, кохлеяобразуван от спираловиден костен канал, canalis spiralis cochleae,който, започвайки от преддверието, се извива като черупка на охлюв, образувайки 2 1/2 кръгови прохода. Костната пръчка, около която са навити проходите на кохлеята, лежи хоризонтално и се нарича модиолус.Спирална костна плоча се простира в кухината на кохлеарния канал през всичките му обороти от modiolus, lamina spiralis ossea.Тази плоча, заедно с кохлеарния канал, разделя кухината на кохлеарния канал на две отделения; вестибюлно стълбище, скала вестибулиобщувайки с вестибюла и scala tympani, scala tympaniкойто се отваря върху скелетираната кост в тъпанчевата кухина през прозореца на кохлеята. Близо до този прозорец в scala tympani има малък вътрешен отвор на акведукта на кохлеята, aqueductus cochleae,чийто външен отвор, apertura externa canaliculi cochleae,лежи на долната повърхност на пирамидата на темпоралната кост.
Мембранният лабиринт, labyrinthus membranaceus, се намира вътре в костния лабиринт и следва повече или по-малко точно очертанията му. Съдържа периферните части на слуховия и гравитационния анализатори. Стените му са изградени от тънка полупрозрачна съединителнотъканна мембрана. Вътре мембранният лабиринт е запълнен бистра течност- ендолимфа. Тъй като мембранният лабиринт е малко по-малък от костния лабиринт, остава празнина между стените на двата - перилимфното пространство, spatium perilymphaticum,изпълнен с перилимфа. В преддверието на костния лабиринт има две части на мембранния лабиринт: утрикулус (елипсовидна торбичка) и сакулус (сферична торбичка). Утрикулусима формата на затворена тръба, заема recessus eilipticus на вестибюла и се свързва отзад с три мембранни полукръгли канала, ductus semicirculares,които лежат в същите костни канали, повтаряйки точно формата на последните. Следователно се разграничават предни, задни и странични мембранни канали, ductus semicircularis anterior, posterior et laterais,със съответните ампули: ampulla membranacea anterior, posterior et laterais. Сакулус- крушовидна торбичка, лежи в recessus sphericus на вестибула и е във връзка с utriculus, както и с дълъг тесен канал, ductus endolymphaticus, който преминава през aqueductus vestibuli и завършва в малко сляпо разширение, saccus endolymphaticus, в дебелината на твърдата обвивка на задната повърхност на пирамидите на темпоралната кост. Малкият тубул, свързващ ендолимфатичния канал с утрикулуса и сакулуса, се нарича ductus utriculosaccularis. С долния си стеснен край, който преминава в тесен ductus reuniens, сакулусът се свързва с мембранозния канал на кохлеята. И дветеВестибуларните торбички са заобиколени от окололимфното пространство.
Мембранният лабиринт в областта на полукръглите канали е окачен върху плътната стена на костния лабиринт чрез сложна система от нишки и мембрани. Това предотвратява изместването на мембранния лабиринт при значителни движения.
Нито перилимфните, нито ендолимфните пространства са затворени за мъртвите. Ендолимфатичното пространство е свързано чрез ендолимфатичния канал с ендолимфатичния сак,
лежи в черепната кухина; това е повече или по-малко еластичен резервоар, който комуникира с вътрешното пространство на полукръглите канали и останалата част от лабиринта.
Структурата на слуховия анализатор.Предната част на мембранозния лабиринт е кохлеарният канал, дуктус кохлеарис,затворен в костната кохлеа, е най-съществената част от органа на слуха. Ductus cochlearis започва със сляп край в recessus cochlearis на вестибула малко по-назад от ductus reuniens, свързвайки кохлеарния канал със сакулуса. След това ductus cochlearis преминава по целия спирален канал на костната кохлеа и завършва сляпо на върха му. В напречно сечение кохлеарният канал има триъгълна форма. Едната от трите му стени се слива с външната стена на костния канал на кохлеята, другата, спирална мембрана,е продължение на костната спирална пластина, простираща се между свободния ръб на последната и външната стена. Третата, много тънка стена на кохлеарния проход, paries vestibuldris ductus cochlearis,опъната наклонено от спираловидната плоча към външната стена.
Membrana spiralis върху базиларната плоча, вградена в нея, lamina basilaris, носи апарат, който възприема звуци - спирален орган.Чрез ductus cochlearis scala vestibuli и scala tympani се отделят една от друга, с изключение на мястото в купола на кохлеята, където има комуникация между тях, наречена кохлеарен отвор, helicotrema. Scala vestibuli комуникира с перилимфатичното пространство на вестибюла, а scala tympani завършва сляпо на прозореца на кохлеята.
Спирален орган, organon spirale,разположени по дължината на целия кохлеарен канал върху базиларната плоча, заемайки най-близката до lamina spiralis ossea част. базиларна плоча, lamina basilaris,се състои от голям брой (24 000) фиброзни влакна с различна дължина, опънати като струни (слухови струни). Според известна теорияХелмхолц (1875), те са резонатори, предизвикващи с вибрациите си възприемането на тонове с различна височина, но според електронната микроскопия тези влакна образуват еластична мрежа, която като цяло резонира със строго степенувани вибрации. Самият спирален орган е изграден от няколко реда епителни клетки, сред които се различават чувствителни слухови клетки с власинки. Той действа като "обратен" микрофон, трансформирайки механичните вибрации в електрически.
Пътища за звук. От функционална гледна точка органът на слуха ( периферна частслух
анализатор) е разделен на две части: 1) звукопроводящ апарат- външно и средно ухо, както и някои елементи (перилимфа и ендолимфа) на вътрешното ухо; 2) апарат за приемане на звук- вътрешно ухо. Въздушните вълни, събрани от ушната мида, се насочват към външния слухов проход, удрят тъпанчето и предизвикват вибрациите му.Вибрация на тъпанчето, степента на напрежението на което се регулира от свиването m. tensor tympani (инервация от n. trigeminus), движи дръжката на чукчето, слято с него. Съответно чукът движи инкуса, а инкусът движи стремето, което се вкарва във fenestra vestibuli, водещо към вътрешното ухо. Степента на изместване на стълбите в прозореца на вестибюла се регулира от свиването m. stapedius (инервация от n. stapedius от n. facialis). По този начин веригата от осикули, свързани подвижно, предава колебателните движения на тимпаничната мембрана, насочени към прозореца на вестибюла.
Движението на стремето навътре в прозореца на вестибюла предизвиква движение на лабиринтната течност, която изпъква мембраната на кохлеарния прозорец навън. Тези движения са необходими за функционирането на високочувствителните елементи на спиралния орган. Перилимфата на вестибюла се движи първа; нейните вибрации по scala vestibuli се изкачват до върха на кохлеята, през helicotrema се предават на перилимфата в scala tympani, по нея се спускат към membrana tympani secundaria, която затваря прозореца на кохлеята, който е слаб точка в костната стена на вътрешното ухо и сякаш се връща в тъпанчевата кухина. От перилимфата звуковите вибрации се предават на ендолимфата, а чрез нея на спиралния орган. По този начин въздушните вибрации във външното и средното ухо, благодарение на системата от слухови осикули на тимпаничната кухина, се превръщат в вибрации на течността на мембранния лабиринт, причинявайки дразнене на специалните слухови космени клетки на спиралния орган, които изграждат рецепторслухов анализатор.
В рецептора, който е като "обратен" микрофон, механичните вибрации на течността (ендолимфа) се преобразуват в електрически вибрации, които характеризират нервен процес, разпространявайки се по проводника към кората на главния мозък. Проводникът на слуховия анализатор се състои от слухови пътища, състоящи се от редица връзки; клетъчното тяло на първия неврон лежи в спиралата на ганглия. Периферният процес на неговите биполярни клетки в спиралния орган започва с рецептори, а централният преминава като част от pars cochlearis. vestibulocochlearis към неговите ядра, nucleus cochlearis dorsalis et ventralis, разположени в областта на ромбовидната ямка. Различни части слухов нервпроизвежда звуци с различни честоти на вибрации.
В тези ядра са разположени телата на вторите неврони, чиито аксони образуват централния слухов път; последният, в областта на задното ядро на трапецовидното тяло, се пресича със същия път на противоположната страна, образувайки страничен контур, lemniscus lateralis. Централно влакно слухов път, идващи от вентралното ядро, образуват трапецовидно тяло и, преминавайки през моста, са част от lemniscus lateralis от противоположната страна. Влакната на централния тракт, излизащи от дорзалното ядро, минават по дъното IVвентрикула под формата на striae medullares ventriculi quarti, проникват в formatio reticularis на моста и заедно с влакната на трапецовидното тяло стават част от страничния контур на противоположната страна. Lemniscus lateralis завършва отчасти в долните коликули на покрива на междинния мозък, отчасти в corpus geniculatum mediale, където се намират третите неврони.
Долните коликули на покрива на средния мозък служат като рефлексен център за слухови импулси. От тях отива в гръбначния мозък tractus tectospinalis, чрез който се осъществяват двигателни реакции към слухови стимули, влизащи в средния мозък. Рефлексните отговори на слуховите импулси могат да бъдат получени и от други междинни слухови ядра - ядрата на трапецовидното тяло и страничния лемнискус, свързани с къси пътища към двигателните ядра на средния мозък, моста и продълговатия мозък.
Завършвайки с образувания, свързани със слуха (долни коликули и corpus geniculatum mediale), слуховите органи и техните колатерали се присъединяват освен това към медиалния надлъжен фасцикулус, чрез който влизат в контакт с ядрата окуломоторни мускулии с моторните ядра на др черепномозъчни нервии гръбначния мозък. Тези връзки обясняват рефлексните реакции на слухови стимули.
Долните коликули на покрива на средния мозък нямат центростремителни връзки с кората. Corpus geniculatum mediale съдържа клетъчните тела на последните неврони, чиито аксони, като част от вътрешната капсула, достигат до кората на темпоралния дял на главния мозък. Кортикален край на слуховия анализаторразположен в gyrus temporalis superior (поле 41). Тук въздушните вълни на външното ухо, които предизвикват движение на слуховите костици в средното ухо и вибрации на течността във вътрешното ухо и се трансформират допълнително в рецептора в нервни импулси, предавани по проводника към кората на главния мозък, са възприемани под формата на звукови усещания. Следователно, благодарение на слуховия анализатор, въздушните вибрации, т.е. обективен феномен на реалния свят, който съществува независимо от нашето съзнание, се отразяват в нашето съзнание под формата на субективно възприемани образи, т.е. звукови усещания.
Това ярък примервалидността на теорията на Ленин за отражението, според която обективно реалният свят се отразява в нашето съзнание под формата на субективни образи. Тази материалистична теория разобличава субективния идеализъм, който, напротив, поставя нашите усещания на първо място.
Благодарение на слуховия анализатор различни звукови стимули, възприемани в нашия мозък под формата на звукови усещания и комплекси от усещания - възприятия, стават сигнали (първите сигнали) на жизненоважни явления на околната среда. Това представлява първата сигнална система на реалността (И. П. Павлов), т.е. конкретно визуално
мислене, характерно и за животните. Човек има способността за абстрактно, абстрактно мислене с помощта на дума, която сигнализира звукови усещания, които са първите сигнали, и следователно е сигнал на сигналите (вторият сигнал). Оттук устна речсъставлява втората сигнална система за действителността, характерна само за човека.
Ушна мида и фиброхрущялни външен слухов канал(с изключение на вътрешния му отдел) получават кръв от клоните на системата на външната каротидна артерия: предната повърхност се доставя от a. auricularis anterior, posterior-a. auricularis posterior, и неговата костна част и вътрешна частхрущялни-а. auricularis profunda (от a. maxillaris interna). Тази артерия отделя клон към външната повърхност на тъпанчето.
Вени, обикновено двойки, следват преминаването на артериалните стволове, те анастомозират с вените на паротидната жлеза, както и с вените на мастоидната област. Вените на външната повърхност на тъпанчето анастомозират с тези на вътрешната повърхност през vv. perforantes.
Лимфни съдовевъншната повърхност на ушната мида и преден отделвъншният слухов канал се простира до лимфните възли, разположени непосредствено пред трагуса и върху паротидната жлеза (както и в нейната дебелина), както и до дълбоките горни цервикални лимфни възли, покрити горна частстерноклеидомастиален мускул.
Лимфни съдовеВътрешната повърхност на ушната мида и задната част на външния слухов канал отиват към лимфните възли на мастоидния процес и отчасти до споменатите дълбоки цервикални горни лимфни възли.
При възпалителни процеси на външното ухоТези лимфни възли често се подуват и понякога дават повод за диференциална диагноза между лимфаденит, от една страна, и заушка или мастоидит, от друга.
Инервация на ушната мида извършен n. auricularis magnus(от горния шиен плексус), n. occipitalis minor (от същия плексус), лицеви и блуждаещи нерви и n. auriculotemporalis (от третия клон на тригеминалния нерв). Според А. М. Талишински, задната повърхност на ушната мида се инервира от големия ушен нерв, малкия тилен (или, по терминологията на този автор, малък тилен-ушен) нерв, задния ушен клон на лицевия нерв и аурикуларен клон на блуждаещия нерв.
Зад ушите областта се инервира от същите нерви, с изключение на скитащия. Инервацията на предната повърхност на ушната мида включва големия ушен нерв и клоновете на аурикулотемпоралния нерв, отиващи към външния слухов канал. Мускулите на ушната мида са обърнати лицев нерв. Външният слухов проход се инервира от ramus auricularis n. vagi (влизащ в прохода през задната му стена) и аурикулотемпоралния нерв, който отделя един клон - n. membranae tympani - към тъпанчето.
Клонове на аурикулотемпоралния нерв, отиващи към външния слухов канал, почти винаги са представени от два ствола (предно-горно и задно-долно), които влизат в слуховия канал на границата на неговите хрущялни и костни участъци. Поради това е препоръчително да се инжектира новокаин в долната задна и предно-горната стена на границата на хрущялните и костните участъци за анестезия на външния слухов канал по време на радикална операция.
нервимножество във външните слоеве на стената на ушния канал. Повечето от тях са без пулпа. Нервни сноповечесто протичат успоредно на кръвоносните съдове и са тясно свързани с жлезите.
Механично дразнене на кожатавъншен слухов канал (например при поставяне на фуния) предизвиква кашличен рефлекс поради дразнене на блуждаещия нерв, а тактилното или калорично дразнене на кожата на дисталната част на външния слухов канал причинява присвиване на очите - ауропалпебралният рефлекс (от тригеминалния нерв към лицевия) или много по-малко постоянния ауролакримален (лакримация).
 Съдържание на темата „Външно ухо. Тъпанчеи кухина":
Съдържание на темата „Външно ухо. Тъпанчеи кухина": Кръвоснабдяването на вътрешното ухо се осъществява от лабиринтната артерия, която в 65% произлиза от предната долна церебеларна артерия, в 29% от базиларната артерия, в 0,5% от задно-долната церебеларна артерия и в 5,5% от различни източници. от дясната и лявата страна (мозъчни и базиларни артерии) (фиг. 4).
Ориз. 4 Артерии на вестибуларния апарат (обозначения на фигурата) [за 17]
Arteria labyrinthi навлиза във вътрешния слухов проход заедно с лицевия и статоакустичния нерв. Лабиринтната артерия е крайна артерия, т.е. няма значителни анастомози с други артерии; изключително рядко дава клонове на долната церебеларна артерия. Ходът на тази артерия най-често е прав (когато изхожда от долната церебеларна артерия) или дъгообразен (когато изхожда от главната артерия). Ширината на лумена на лабиринтната артерия е малка и може да се класифицира като субмилиметрична. На входа на вътрешното ухо лабиринтната артерия се разделя на предна вестибуларна артерия и обща кохлеарна артерия, която завършва с разделение на вестибулокохлеарна и кохлеарна артерия. Предната вестибуларна артерия доставя горните части на вестибуларния лабиринт, включително хоризонталния полукръгъл канал, utriculus macula и вестибуларен нерв. Общата кохлеарна артерия доставя долните части на вестибуларния лабиринт и кохлеята. Между тези клонове на лабиринтната артерия на нивото на горните части на лабиринта почти няма анастомози, за разлика от наличието на колатерали на нивото на долните части на лабиринта.
Тези анатомични особености определят различната чувствителност на лабиринтните участъци към исхемия. Чувствителността към исхемия на мембранозния лабиринт като орган на баланса и слуха се дължи и на факта, че няма колатерално кръвообращение от съдовете на ушната капсула.
Ушният лабиринт е най-чувствителен към развитието на исхемични състояния във вертебробазиларната област. Замаяността при тези условия се причинява от разликата между кръвния поток през дясната и лявата лабиринтна артерия или по-големите съдове на вертебробазиларната система и следователно разликата в кръвоснабдяването на десния и левия лабиринт.
Вестибуларните ядра заемат значителна площ в страничните части на мозъчния ствол и се кръвоснабдяват от проникващи клонове от вертебралните и базиларните артерии. Клинично важно, тази област е особено податлива както на исхемично, така и на хеморагично увреждане.
Вестибуларната система изпълнява три основни функции (фиг. 1): пространствена ориентация, контрол на баланса и стабилизиране на образа.
3.1 Ориентация в пространството
Функцията за пространствена ориентация е изключително важна - необходимо условиеза контролиране на стойката на тялото, движението и взаимодействието с заобикаляща среда. За оптималното изпълнение на тази функция е необходимо да се получава информация от всички сетива. Вестибуларният апарат е част сложна системасензорна интерпретация и интеграция. Визуалното наблюдение на нашата позиция в околната среда помага да се определи абсолютната позиция. Пациентите с патология на вестибуларния апарат компенсират до голяма степен тази патология със зрителна информация. Несъответствието във функционирането на вестибуларната, визуалната и проприоцептивната система и липсата на синхронна аферентация към центровете причиняват развитието на нестабилност.
3.2 Стабилизация на изображението
Изображение, което се движи през ретината по-бързо от 2-3 градуса в секунда, не може да бъде обработено от зрителната система без замъгляване на изображението. Поради тази причина движещото се изображение трябва да бъде стабилизирано в ретината. Ако целта се движи, тогава окуломоторната система е в състояние да премести погледа, което му позволява, благодарение на оптокинетичния рефлекс, да следва целта. Ако целта е неподвижна, изображението на ретината на целта също ще се премести, ако индивидът направи движение на главата. В тази ситуация стабилизирането на изображението се постига чрез движение на очите в посока, обратна на тази, в която се движи главата (компенсаторно движение на очите или вестибуло-окуларен рефлекс).
3. 2.1 Сакадичен поглед
Окуломоторната реакция под формата на рязко движение на двете очи към обекта на интерес се нарича сакада (от френски: saccade - внезапно спиране на кон с рязко движение). Изображението, появяващо се в периферното зрително поле, бързо се премества в централната ямка (областта с най-голяма пространствена разделителна способност) за подробен анализ чрез бързо потрепване (потрепване) на очите, по време на което зрението се потиска за кратко. Прецизността на тези движения се регулира постоянно чрез визуална обратна връзка. В този случай стимулът от мозъчната кора достига до ядрото на абдуценсния нерв от противоположната страна и, след пресичане в горните части на моста, до ипсилатералното ядро на окуломоторния нерв. Това води до едновременно свиване на латералния ректус мускул на едното око и респективно медиалния ректус мускул на противоположното око и като следствие до благоприятна ротация на очните ябълки. Такъв хармоничен нервен механизъм е възможен благодарение на синхронната работа на влакната в медиалния надлъжен фасцикулус.
3.2.2 Проследяване на движенията на очите
Окуломоторната система също е в състояние да следва целта, докато се движи. Рефлексът, включен в този процес, се нарича гладко преследване. Рефлексът на зрителното преследване контролира процесите по пътя от централната фовеа на ретината през страничното геникулатно тяло (corpus geniculatum laterale) в таламуса (визуален таламус) до зрителната зона на мозъчната кора (19-то поле на париеталната- тилната област). В резултат на това двигателна команда се изпраща през кортико-тектални и кортико-тегментални влакна към средния мозък и моста, малкия мозък и вестибуларните ядра към окуломоторното ядро и наклонените (екстраокулярни) окуломоторни мускули. Времето на забавяне е 70 милисекунди. По време на плавно проследяване движенията трябва да бъдат много точни, тъй като фовеята на ретината заема площ от само 1 дъгова степен - изображението на движещ се обект може лесно да се изплъзне от тази област. За офталмологично здрав човек скоростта на движение на стимула през ретината не трябва да надвишава 30–60 градуса в секунда. При по-високи скорости този механизъм става неадекватен и са необходими коригиращи сакади за фиксиране на целта в централната фовеа.
3.2.3. Оптокинетичен рефлекс
В кората на мозъка оптокинетичният рефлекс следва същия път като рефлекса на гладко преследване, но използва информация, получена от цялата ретина. Например, когато гледаме минаващ влак, изображението на влака се движи през ретината и зрителната система изчислява скоростта на изображението в зрителната кора. Въз основа на тази информация се генерират сдвоени (конюгирани) движения на очите (оптокинетичен нистагъм) със скорост, която съответства на скоростта на движение на целта. Инициирането на бавния компонент на оптокинетичния нистагъм се определя от преминаването на отражения по периферната ретина. Бързият компонент на нистагъм играе по-активна роля с участието на по-високи кортикални центрове, свързани с фиксиращия рефлекс. Подкорковият оптокинетичен рефлекс позволява на бебетата да стабилизират визуални образи, движещи се през ретината. През първите месеци от живота зрението е слабо развито; Няма възможност за плавно проследяване на движението на обект и изглежда, че бебетата възприемат само големи обекти, които привличат вниманието. Скоростта на движение на изображението се изчислява от всяко око поотделно в двете ядра на оптичния тракт чрез паметта за съхраняване на стойности на скоростта (разположена в ядрото пред ядрото на хипоглосния нерв и малкия мозък) и в зависимост по данни за това се активират косите (екстраокуларни) екстраокуларни мускули. Този път е активен и при възрастни, когато възникне подсъзнателно наблюдение на цел. Подкорковият път започва да функционира от момента на раждането, заобикаляйки центъра на погледа ретикуларна формацияМостът, който отговаря за координацията на движенията на двете очи. Следователно при новородени оптокинетичните рефлекси се появяват във всяко око независимо, докато се развие бинокулярно зрение, в което участва кората на главния мозък. Формирането на кортикалния оптокинетичен рефлекс отнема време. Преди очите да започнат да се движат, паметта за съхранение на скорост трябва да е пълна. Остатъчната активност, съхранена в паметта, е отговорна за движението на очите (нистагъм), което възниква, когато визуален стимул внезапно изчезне. Този феномен се нарича "оптокинематичен след нистагъм" (OKAN) и често се използва, за да се определи дали функцията за съхранение на скоростта е увредена. Дисфункция на системата за съхранение на информация за скоростта или намаляване на количеството информация, получена от лабиринтите (повреда на лабиринта) обикновено води до съкратени реакции след въртене. Пасивен отговор на оптокинетичен стимул може да се получи с помощта на оптокинетичен барабан.
3.2.4 Окуловестибуларни рефлекси
Изображението на целта върху ретината също ще се движи, когато индивидът движи главата си, въпреки че целта може да е неподвижна. Рефлексите, отговорни за движението на очите по време на зрение, обикновено са твърде бавни, за да стабилизират изображението на ретината, ако главата се движи бързо. Окуловестибуларните рефлекси са доста бърз механизъм, докато движенията на очите в съответствие със скоростта се причиняват директно от стимулация на лабиринтите. Този рефлекс е може би най-бързият в тялото, с време на забавяне от 7-10 милисекунди (от които 2 милисекунди се изразходват за механичния процес, задвижван от пречупването на светлината, и около 5 милисекунди за провеждането и свиването на нервния импулс). / релаксация на косите (извъночните) извъночни мускули).
От лабиринтите информацията се предава към вестибуларното ядро на мозъчния ствол и след това към окуломоторното ядро, предизвиквайки компенсаторни движения на очите. В резултат на това скоростта на движение на изображението през ретината се свежда до минимум чрез движение на очите в посока, обратна на тази, в която се движи главата. След това възниква визуална обратна връзка: зрителният кортекс обработва информация за оставащото движение на изображението и изпраща сигнал до вестибуларното ядро през центровете за поглед в моста и малкия мозък, за да регулира силата на рефлекса. В лабораторни условия силата на рефлекса се регулира в диапазона до 30% в рамките на няколко минути.
3.3 Възприятие.
Различни зони в париеталната и темпоралната кора се активират от стимули, излъчвани от лабиринтите, зрителните органи и проприорецепторите. Предполага се, че тези мултисензорни области участват в пространствената ориентация и възприемането на движението. Поради тази причина функцията на вестибуларния кортекс изглежда е разпределена между няколко мултисензорни области и интегрирана в по-голяма мрежа за „пространствено внимание“ и сензомоторно управление. Вестибуларната зона на парието-инсуларната област на кората на главния мозък се счита за основна зона във вестибуларната система на кората. Тя е представена от двете страни, като дясното полукълбо играе доминираща роля.
КОНТРОЛНИ ВЪПРОСИ:
1. Какво определя разнообразието от вестибуларни реакции, които се развиват при представяне на надпрагов стимул.
2. Кой е адекватен стимул за ампуларния и отолитния апарат.
3. На какво се дължи различната чувствителност на частите на ушния лабиринт към исхемия.
4. От кой артериален басейн се захранва вътрешното ухо?
5. Какво причинява бързите и бавните компоненти на оптокинетичния нистагъм.
Инервация на ушната мидауникален (фиг. 2, 3). Въпреки факта, че ушната мида е лишена от всякакви специализирани функции, тя съдържа аферентни нерви от соматичен и висцерален произход.
Първите са представени от клоните на два големи нерва - тригеминалния ( н. тригеминус) и цервикалния плексус ( plexus cervicalis), вторият - от клоните на вагусния нерв ( н. вагус), глософарингеален нерв ( н. glossopharyngeus) и лицевия нерв ( н. фациалис).

Система на тригеминалния нерв. Аурикулотемпорален нерв ( н. auriculotemporalis), най-големият, клон мандибуларен нерв- третият клон на тригеминалния нерв на нивото на ухото дава нерви на външния слухов канал ( nn. meatus acustici externi), които под формата на два крайни клона проникват в стената на външния слухов канал, инервират началото на външния слухов канал, совалката на раковината и корена на спиралата.

Предни ушни нерви ( nn. auricularis anteriores) са основните нерви на ушната мида, принадлежащи към тригеминалната система. Тяхната инервационна зона: трагус, част от спиралата, триъгълна ямка, антихеликс, част от скафоида, горна половина на лобула на ушната мида (ушна мида). Границите на инервацията на ушната мида от тригеминалния нерв са показани на фиг. 4.
Нервна система на цервикалния плексус. Голям ушен нерв ( н. auricularis magnus) започва от третия и по-рядко четвъртия цервикален нерв (С 3 -С 4). На долния полюс на ушната мида се разделя на преден и заден клон. Преден клон ( r. преден) инервира вътрешна повърхностаурикула, след което, преминавайки през него, излиза на външна повърхности инервира по-голямата част от ушната мида, част от спиралата, скафоидната кост, спираловидната бразда, антихеликса, триъгълната ямка и краищата на кухината на раковината. По този начин дъното на кухината на раковината остава извън сферата на инервация на предния клон.
Заден клон ( r. заден) се разклонява главно в кожата на вътрешната повърхност на ухото, частично преминава към външната повърхност и инервира част от спиралата, антихеликса и триъгълната ямка.
Малък тилен нерв ( н. occipitalis minor), произхождащ от втория и третия нерв на цервикалния плексус (C 2 -C 3), горния клон ( r. превъзхождащ) инервира горния полюс на вътрешната повърхност на ухото, част от спиралата и триъгълната ямка.
Долен клон ( r. непълноценен) също инервира част от вътрешната повърхност на ухото и, движейки се по спиралата към външната повърхност, инервира част от спиралата и топа.
Важно е да се отбележи, че малкият тилен нерв има анастомози с големия ушен или лицевия нерв. Освен това, използвайки свързващи клонове ( rr. комуниканти) нервите на цервикалния плексус се свързват с цервикалния симпатикови възли (ganglion sympaticus). Това обстоятелство е изключително ценно, тъй като ефектите върху акупунктурните точки, разположени в зоната на инервация на цервикалните нерви, могат да бъдат предадени на симпатиковите ганглии и след това да се разпространят по цялата симпатикова верига.

Границите на инервацията на ушната мида от нервите на цервикалния плексус са показани на фиг. 5.
Система на лицевия нерв. Според данните на V.P. Vorobyov, аурикулотемпоралният нерв (система на тригеминалния нерв) на нивото на ухото е свързан с клоновете на междинния нерв на Wriesberg (система на лицевия нерв) чрез анастоматични клонове ( rr. anastomotici cum n. faciale), които инервират средната част (проекцията на кухината на раковината) на вътрешната повърхност на ухото, входа на задната стена на външния слухов канал, областта на трагуса и костите, кухината на раковината, триъгълната ямка, антихеликса , по-голямата част от ушната мида (ушната мида). Границите на инервацията на ушната мида от лицевия нерв са показани на фиг. 6.

Системата на блуждаещия и глософарингеалния нерв. На нивото на югуларния отвор на черепа отделен ушен клон се отклонява от ствола на блуждаещия нерв ( r. auricularis n. vagi), който заедно с клона на глософарингеалния нерв, който веднага се присъедини ( r. глософарингеален нерв) се насочва към ушната мида югуларна вена, през дебелината на пирамидата на темпоралната кост.

След като напуснат костния канал, тези нерви се разделят на два клона, които инервират вътрешната повърхност на ухото, субтрагалната област, тристранната ямка и ушната мида. Границите на инервацията на ушната мида от вагуса и глософарингеален нервса представени на фиг. 7 и 8.


Римляните и древните гърци много обичали октоподите и постоянно ги ловували. Любовта е минала през вековете и сега...

Октоподът не изисква дълго варене. Малките замразени октоподи се нуждаят само от 5 минути варене във вряща вода. цял...

Получавам много въпроси за агар-агар по имейл, затова реших да напиша още една статия за този уникален...

Докато чете езана, ръцете на мюезина трябва да докосват ушните му миди и погледът му трябва да бъде насочен към Кааба. След...

През 2018 г. Рождественският пост ще започне на 28 ноември. През този период православните вярващи се подготвят да празнуват Коледа...

Създаването на семейство е мечтата на повечето жени. Те искат да имат любящ съпруг и куп деца. Но не винаги е връзка...

Тази статия съдържа: най-силната молитва за развод - информация взета от цял свят, електронна...

Информационен сайт за икони, молитви, православни традиции. Молитва за скандали и кавги в семейството, със съпруга, с...

Каква би била Нова година без шампанско, мандарини, оливие, заливка и любимата на всички „Херинга под шуба“. Тук с...

Нека подготвим необходимите съставки за бисквитките. Първото нещо, което трябва да направите, е да сложите водата да заври. Нас...

Възможно ли е записване на служител за длъжността финансов директор - главен счетоводител? Главният счетоводител твърди...

Ръководителят на малък бизнес може лесно да управлява бюджета самостоятелно. ПРОВЕРЕНО! Ако успееш...

Създаването на нови проекти е свързано с предварителна икономическа обосновка за тяхната осъществимост, последващо...
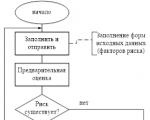
Отчетността се генерира от RM, съгласува се (одобрява) от Комитета по риска към Управителния съвет и се предава на...

Октоподът не изисква дълго варене. Малките замразени октоподи се нуждаят само от 5 минути варене...

Получавам много въпроси за агар-агар по имейл, затова реших да напиша друга статия за него...