The most complete interpretation of sleep orange
Orange juice. The symbolic meaning of orange juice in dream books is pleasure and temptation. Quite often we...
Changes in the transport of water and ions in the renal tubules occur under the influence of nervous stimuli or hormones. The essence of the diverse effects of these substances can be reduced to two main methods of intracellular reactions. An example of one is the action of aldosterone, the other is ADH. Aldosterone, once introduced into the blood, increases sodium reabsorption after a fairly long period latent period, equal to 45-120 minutes.
This hormone enters the cell through the peritubular plasma membrane and binds to a stereospecific protein in the cytoplasm. This process takes 30-45 minutes, after which the aldosterone-receptor complex is transferred to the nucleus and interacts with the acceptor regions of nuclear chromatin. This causes gene transcription - activation of a section of DNA promotes the synthesis of messenger RNA, which moves from the nucleus to the cytoplasm and stimulates the synthesis of a new protein in the ribosomes. Its relative molecular weight is about 12,000. There are a number of hypotheses about the point of application in the cell of this protein and the mechanism by which it increases sodium transport.
One hypothesis is that this protein is a component of a transporter or permease, facilitating the entry of sodium into the cell through the apical membrane. Another hypothesis attaches leading importance to the activation of the pump - Na, K-ATPase. Finally, according to the metabolic hypothesis, this protein serves as a component of the cell's energy metabolism system; it enhances mitochondrial ATP synthesis. It is also suggested that this protein may act on not one, but two components of the system - the penetration and active removal of sodium from the cell, which automatically increases energy consumption and leads to an increase in the formation of ATP. A fairly in-depth study of the mechanism of action of aldosterone has essential for the clinic, the mode of action, the reason for the long latent period and the essence of the influence of diuretics - opironolactones - competitive aldosterone antagonists (veroshpiron, aldactone) become clear. These substances inhibit the binding of aldosterone to receptor proteins located in the cytoplasm and nucleus.
It has been established that the main site of action of aldosterone in the nephron is the most terminal sections of the distal convoluted tubule and primary departments collecting ducts. Spironolactones also act in these same cells. The above data on the mechanism of cellular influence of aldosterone relate only to its effect on sodium reabsorption; The hormone-induced increase in potassium secretion is not mediated through the genetic apparatus of the cell.
A different mechanism of intracellular action is characteristic of ADH.
On outer surface The receptor protein for ADH is localized in the basal plasma membrane of collecting duct cells. Composite element membrane receptor is the enzyme adenylate cyclase, which, upon activation of the receptor, catalyzes inside membrane formation of cAMP 3´, 5´-AMP from ATP. The exact mechanism of all subsequent stages of action of this substance in the cell of the collecting duct is not yet clear.
It is known that 3",5"-AMP activates the transfer of high-energy phosphate using the enzyme protein phosphase on the series or threonine protein phosphatase. This enzyme is located on the inside of the apical membrane of the cell. These reactions ultimately cause an increase in the permeability of the tubular wall to water. 3´,5"-AMP, which induced the entire sequence of intracellular reactions, is converted into physiologically inactive 5"-AMP with the help of the enzyme cyclic nucleotide phosphodiesterase.
It is significant that the inhibitor of this enzyme is the substance theophylline, which is very often used in the clinic. A similar sequence of intracellular reactions is characteristic of the action of catecholamines and a number of peptide hormones, such as PG, on the nephron cell. The difference lies in the specificity of the receptor, located on the outside of the basal plasma membrane, and another final link in intracellular reactions initiated by 3,5"-AMP.
The pathways for water reabsorption are still a matter of debate. According to one hypothesis, the final stage of the action of 3,5"-AMP is an increase in the permeability for water of the apical plasma membrane, and water, crossing the cell, moves along the osmotic gradient into the intercellular fluid and blood. According to another hypothesis, under the influence of ADH with the participation of 3,5"-AMP, secretion of hyaluronidase occurs. A.G. Ginetsinsky (1963) believed that this enzyme, by depolymerizing glycosaminoglycans of the intercellular substance, increases the permeability of the membrane to water.
The approaches to the series described above pathological conditions made it possible to identify their significant significance for understanding the pathogenesis of nephrogenic diabetes insipidus, impaired concentrating ability of the kidney in chronic renal failure, etc. Thus, in one of the forms of nephrogenic diabetes insipidus, a decrease in the secretion of hyaluronidase and the formation of 3", 5"-AMP under the influence of ADH was found.
Clinical Nephrology
edited by EAT. Tareeva
There are three possible options mechanism of action of hormones.
Membrane or local mechanism- lies in the fact that the hormone at the site of binding to the cell membrane changes its permeability to metabolites, for example, glucose, amino acids, and some ions. The supply of glucose and amino acids, in turn, affects the biochemical processes in the cell, and changes in the distribution of ions on both sides of the membrane affect the electrical potential and function of the cells. The membrane type of hormone action is rarely found in isolated form. For example, insulin has both a membrane (causes local changes in the transport of ions, glucose and amino acids) and a membrane-intracellular type of action.
Membrane-intracellular the type of action (or indirect) is characteristic of hormones that do not penetrate the cell and therefore affect metabolism through an intracellular chemical messenger, which is the authorized representative of the hormone inside the cell. The hormone affects function through membrane receptors signaling systems(usually these are enzymes) that trigger the formation or entry of intracellular mediators. And the latter, in turn, affect the activity and quantity of various enzymes and thereby change the metabolism in the cell.
Cytosolic mechanism action is characteristic of lipophilic hormones that are able to penetrate through the lipid layer of the membrane into the cell, where they enter into a complex with cytosolic receptors. This complex regulates the number of enzymes in the cell, selectively influencing the activity of nuclear chromosome genes, and thereby changing the metabolism and functions of the cell. This type of hormone action is called direct, in contrast to the membrane-intracellular action, when the hormone regulates metabolism only indirectly, through intracellular intermediaries.
Hormones of the thyroid and parathyroid glands
Hormones thyroid gland
The thyroid gland secretes two groups of hormones: different influence on metabolism. The first group is iodothyronines: thyroxine and triiodothyronine. These hormones regulate energy metabolism and influence cell division and differentiation, determining the development of the organism. Iodothyronines act on many tissues of the body, but in to the greatest extent on the tissue of the liver, heart, kidneys, skeletal muscles and to a lesser extent on adipose and nervous tissue.
With hyperfunction of the thyroid gland (hyperthyroidism), excessive formation of iodothyronines is observed. A characteristic feature thyrotoxicosis is the accelerated breakdown of carbohydrates and fats (mobilized from fat depots). The rapid combustion of fatty acids, glycerol and glycolysis products requires a large consumption of oxygen. Mitochondria increase in size, swell, and their shape changes. Therefore, thyrotoxicosis is sometimes called “mitochondrial disease.” Externally, hyperthyroidism manifests itself in the form of the following symptoms: increased basal metabolism, increased body temperature (increased heat production), weight loss, severe tachycardia, increased nervous excitability, bulging eyes, etc. These disorders can be relieved either by surgical removal of part of the thyroid gland, or with the help of drugs that suppress it activity.
With hypofunction (hypothyroidism) of the thyroid gland there is a lack of iodothyronines. Hypothyroidism in early childhood called cretinism or myxedema in children, and in adults simply myxedema. Cretinism is characterized by pronounced physical and mental retardation. This is explained by a decrease in the effect of iodothyronines on cell division and differentiation, which entails slow and abnormal growth bone tissue, impaired differentiation of neurons. In adults, myxedema manifests itself in a decrease in basal metabolism and body temperature, memory impairment, and impaired skin(dryness, peeling), etc. In the tissues of the body, the metabolism of carbohydrates and fats and all energy processes are reduced. Hypothyroidism is eliminated by treatment with iodothyronine drugs.
The second group includes calciotonin (a protein with a molecular weight of 30,000), it regulates phosphorus-calcium metabolism, its action is discussed below.
 4 main metabolic regulation systems: Central nervous system(due to signal transmission via nerve impulses and neurotransmitters); Endocrine system(with the help of hormones that are synthesized in the glands and transported to target cells (in Fig. A); Paracrine and autocrine systems (with the participation of signaling molecules secreted from cells into the intercellular space - eicosanoids, histamines, gastrointestinal hormones, cytokines) (on Fig. B and C); The immune system(through specific proteins - antibodies, T-receptors, histocompatibility complex proteins.) All levels of regulation are integrated and act as a single whole.
4 main metabolic regulation systems: Central nervous system(due to signal transmission via nerve impulses and neurotransmitters); Endocrine system(with the help of hormones that are synthesized in the glands and transported to target cells (in Fig. A); Paracrine and autocrine systems (with the participation of signaling molecules secreted from cells into the intercellular space - eicosanoids, histamines, gastrointestinal hormones, cytokines) (on Fig. B and C); The immune system(through specific proteins - antibodies, T-receptors, histocompatibility complex proteins.) All levels of regulation are integrated and act as a single whole.
 The endocrine system regulates metabolism through hormones. Hormones (ancient Greek ὁρμάω - I excite, encourage) - - biologically active organic compounds, which are produced in small quantities in the endocrine glands, carry out humoral regulation of metabolism and have a different chemical structure.
The endocrine system regulates metabolism through hormones. Hormones (ancient Greek ὁρμάω - I excite, encourage) - - biologically active organic compounds, which are produced in small quantities in the endocrine glands, carry out humoral regulation of metabolism and have a different chemical structure.
 Classic hormones have a number of characteristics: Distance of action - synthesis in the endocrine glands, and regulation of distant tissues Selectivity of action Strict specificity of action Short duration of action They act in very low concentrations, under the control of the central nervous system and regulation of their action is carried out in most cases according to the type feedback Act indirectly through protein receptors and enzymatic systems
Classic hormones have a number of characteristics: Distance of action - synthesis in the endocrine glands, and regulation of distant tissues Selectivity of action Strict specificity of action Short duration of action They act in very low concentrations, under the control of the central nervous system and regulation of their action is carried out in most cases according to the type feedback Act indirectly through protein receptors and enzymatic systems
 Organization of neurohormonal regulation There is a strict hierarchy or subordination of hormones. Maintaining hormone levels in the body in most cases provides a negative feedback mechanism.
Organization of neurohormonal regulation There is a strict hierarchy or subordination of hormones. Maintaining hormone levels in the body in most cases provides a negative feedback mechanism.


 Regulation of hormone levels in the body Changing the concentration of metabolites in target cells through a negative feedback mechanism suppresses the synthesis of hormones, acting either on endocrine glands, or to the hypothalamus. There are endocrine glands for which there is no regulation by tropic hormones - a couple thyroid, adrenal medulla, renin-aldosterone system and pancreas. They are controlled by nervous influences or concentration certain substances in blood.
Regulation of hormone levels in the body Changing the concentration of metabolites in target cells through a negative feedback mechanism suppresses the synthesis of hormones, acting either on endocrine glands, or to the hypothalamus. There are endocrine glands for which there is no regulation by tropic hormones - a couple thyroid, adrenal medulla, renin-aldosterone system and pancreas. They are controlled by nervous influences or concentration certain substances in blood.
 Classification of hormones by biological functions; by mechanism of action; By chemical structure; There are 4 groups: 1. Protein-peptide 2. Amino acid-derived hormones 3. Steroid hormones 4. Eicosanoids
Classification of hormones by biological functions; by mechanism of action; By chemical structure; There are 4 groups: 1. Protein-peptide 2. Amino acid-derived hormones 3. Steroid hormones 4. Eicosanoids


 1. Protein - peptide hormones Hormones of the hypothalamus; pituitary hormones; pancreatic hormones - insulin, glucagon; thyroid hormones and parathyroid glands– calcitonin and parathyroid hormone, respectively. They are produced mainly by targeted proteolysis. Hormones have a short lifespan and have from 3 to 250 AMK residues.
1. Protein - peptide hormones Hormones of the hypothalamus; pituitary hormones; pancreatic hormones - insulin, glucagon; thyroid hormones and parathyroid glands– calcitonin and parathyroid hormone, respectively. They are produced mainly by targeted proteolysis. Hormones have a short lifespan and have from 3 to 250 AMK residues.
 The main anabolic hormone is insulin, the main catabolic hormone is glucagon.
The main anabolic hormone is insulin, the main catabolic hormone is glucagon.
 Some representatives of protein-peptide hormones: thyroliberin (pyroglu-his-pro-NN HH 22), insulin and somatostatin.
Some representatives of protein-peptide hormones: thyroliberin (pyroglu-his-pro-NN HH 22), insulin and somatostatin.
 2. Hormones are derivatives of amino acids. They are derivatives of the amino acid tyrosine. These include the thyroid hormones - triiodothyronine (II 33) and thyroxine (II 44), as well as adrenaline and norepinephrine - catecholamines.
2. Hormones are derivatives of amino acids. They are derivatives of the amino acid tyrosine. These include the thyroid hormones - triiodothyronine (II 33) and thyroxine (II 44), as well as adrenaline and norepinephrine - catecholamines.


 3. Hormones of steroid nature Synthesized from cholesterol (in Fig.) Hormones of the adrenal cortex - corticosteroids (cortisol, corticosterone) Hormones of the adrenal cortex - mineralocorticoids (andosterone) Sex hormones: androgens (19 “C”) and estrogens (18 “C” )
3. Hormones of steroid nature Synthesized from cholesterol (in Fig.) Hormones of the adrenal cortex - corticosteroids (cortisol, corticosterone) Hormones of the adrenal cortex - mineralocorticoids (andosterone) Sex hormones: androgens (19 “C”) and estrogens (18 “C” )

 Eicosanoids The precursor of all eicosanoids is arachidonic acid. They are divided into 3 groups - prostaglandins, leukotrienes, thromboxanes. Eikazonoids are mediators (local hormones) - a widespread group of signaling substances that are formed in almost all cells of the body and have a short range of action. This is how they differ from classical hormones synthesized in special cells of the endocrine glands. .
Eicosanoids The precursor of all eicosanoids is arachidonic acid. They are divided into 3 groups - prostaglandins, leukotrienes, thromboxanes. Eikazonoids are mediators (local hormones) - a widespread group of signaling substances that are formed in almost all cells of the body and have a short range of action. This is how they differ from classical hormones synthesized in special cells of the endocrine glands. .
 Characteristic different groups eikasonoids Prostaglandins (Pg) - are synthesized in almost all cells, except erythrocytes and lymphocytes. The following types of prostaglandins are distinguished: A, B, C, D, E, F. The functions of prostaglandins are reduced to changing the tone of the smooth muscles of the bronchi, genitourinary and vascular systems, gastrointestinal tract, while the direction of changes varies depending on the type of prostaglandins and conditions. They also affect body temperature. Prostacyclins are a subtype of prostaglandins (Pg I), but additionally have a special function - they inhibit platelet aggregation and cause vasodilation. They are especially actively synthesized in the endothelium of the vessels of the myocardium, uterus, and gastric mucosa. .
Characteristic different groups eikasonoids Prostaglandins (Pg) - are synthesized in almost all cells, except erythrocytes and lymphocytes. The following types of prostaglandins are distinguished: A, B, C, D, E, F. The functions of prostaglandins are reduced to changing the tone of the smooth muscles of the bronchi, genitourinary and vascular systems, gastrointestinal tract, while the direction of changes varies depending on the type of prostaglandins and conditions. They also affect body temperature. Prostacyclins are a subtype of prostaglandins (Pg I), but additionally have a special function - they inhibit platelet aggregation and cause vasodilation. They are especially actively synthesized in the endothelium of the vessels of the myocardium, uterus, and gastric mucosa. .
 Thromboxanes and leukotrienes Thromboxanes (Tx) are formed in platelets, stimulate their aggregation and cause constriction of small vessels. Leukotrienes (Lt) are actively synthesized in leukocytes, in the cells of the lungs, spleen, brain, and heart. There are 6 types of leukotrienes: A, B, C, D, E, F. In leukocytes, they stimulate motility, chemotaxis and migration of cells to the site of inflammation. They also cause contraction of the bronchial muscles in doses 100-1000 times less than histamine.
Thromboxanes and leukotrienes Thromboxanes (Tx) are formed in platelets, stimulate their aggregation and cause constriction of small vessels. Leukotrienes (Lt) are actively synthesized in leukocytes, in the cells of the lungs, spleen, brain, and heart. There are 6 types of leukotrienes: A, B, C, D, E, F. In leukocytes, they stimulate motility, chemotaxis and migration of cells to the site of inflammation. They also cause contraction of the bronchial muscles in doses 100-1000 times less than histamine.
 Interaction of hormones with receptors of target cells To exhibit biological activity, the binding of hormones to receptors must lead to the formation of a signal that causes a biological response. For example: the thyroid gland is a target for thyrotropin, under the influence of which the number of acinar cells increases and the rate of synthesis of thyroid hormones increases. Target cells distinguish the corresponding hormone due to the presence of the corresponding receptor.
Interaction of hormones with receptors of target cells To exhibit biological activity, the binding of hormones to receptors must lead to the formation of a signal that causes a biological response. For example: the thyroid gland is a target for thyrotropin, under the influence of which the number of acinar cells increases and the rate of synthesis of thyroid hormones increases. Target cells distinguish the corresponding hormone due to the presence of the corresponding receptor.

 General characteristics of receptors Receptors can be located: - on the surface cell membrane- inside the cell - in the cytosol or in the nucleus. Receptors are proteins that can consist of several domains. Membrane receptors have a hormone recognition and binding domain, a transmembrane and a cytoplasmic domain. Intracellular (nuclear) – hormone binding domains, DNA and protein binding domains that regulate transduction.
General characteristics of receptors Receptors can be located: - on the surface cell membrane- inside the cell - in the cytosol or in the nucleus. Receptors are proteins that can consist of several domains. Membrane receptors have a hormone recognition and binding domain, a transmembrane and a cytoplasmic domain. Intracellular (nuclear) – hormone binding domains, DNA and protein binding domains that regulate transduction.
 The main stages of hormonal signal transmission: through membrane (hydrophobic) and intracellular (hydrophilic) receptors. These are the fast and slow ways.
The main stages of hormonal signal transmission: through membrane (hydrophobic) and intracellular (hydrophilic) receptors. These are the fast and slow ways.
 The hormonal signal changes the rate of metabolic processes by: - changing the activity of enzymes - changing the number of enzymes. According to the mechanism of action, hormones are distinguished: - interacting with membrane receptors (peptide hormones, adrenaline, eicosanoids) and - interacting with intracellular receptors (steroid and thyroid hormones)
The hormonal signal changes the rate of metabolic processes by: - changing the activity of enzymes - changing the number of enzymes. According to the mechanism of action, hormones are distinguished: - interacting with membrane receptors (peptide hormones, adrenaline, eicosanoids) and - interacting with intracellular receptors (steroid and thyroid hormones)
 Transmission of hormonal signals through intracellular receptors for steroid hormones (adrenocortical hormones and sex hormones), thyroid hormones (T 3 and T 4). Slow transmission type.
Transmission of hormonal signals through intracellular receptors for steroid hormones (adrenocortical hormones and sex hormones), thyroid hormones (T 3 and T 4). Slow transmission type.

 Transmission of a hormonal signal through membrane receptors The transmission of information from the primary messenger of the hormone occurs through the receptor. The receptors transform this signal into a change in the concentration of secondary messengers, called second messengers. The coupling of the receptor with the effector system is carried out through the GG protein. General mechanism, through which biological effects are realized is the process of “phosphorylation - dephosphorylation of enzymes.” There are different mechanisms for transmitting hormonal signals through membrane receptors - adenylate cyclase, guanylate cyclase, inositol phosphate systems and others.
Transmission of a hormonal signal through membrane receptors The transmission of information from the primary messenger of the hormone occurs through the receptor. The receptors transform this signal into a change in the concentration of secondary messengers, called second messengers. The coupling of the receptor with the effector system is carried out through the GG protein. General mechanism, through which biological effects are realized is the process of “phosphorylation - dephosphorylation of enzymes.” There are different mechanisms for transmitting hormonal signals through membrane receptors - adenylate cyclase, guanylate cyclase, inositol phosphate systems and others.
 The signal from the hormone is transformed into a change in the concentration of secondary messengers - c. AMF, c. GTP, IF 3, DAG, CA 2+, NO.
The signal from the hormone is transformed into a change in the concentration of secondary messengers - c. AMF, c. GTP, IF 3, DAG, CA 2+, NO.
 The most common system for transmitting hormonal signals through membrane receptors is the adenylate cyclase system. The hormone-receptor complex is associated with a G protein, which has 3 subunits (α, β and γ). In the absence of the hormone, the α subunit is associated with GTP and adenylate cyclase. The hormone-receptor complex leads to the cleavage of the βγ dimer from α GTP. The α subunit of GTP activates adenylate cyclase, which catalyzes the formation of cyclic AMP (c. AMP). c. AMP activates protein kinase A (PKA), which phosphorylates enzymes that change the rate of metabolic processes. Protein kinases are classified as A, B, C, etc.
The most common system for transmitting hormonal signals through membrane receptors is the adenylate cyclase system. The hormone-receptor complex is associated with a G protein, which has 3 subunits (α, β and γ). In the absence of the hormone, the α subunit is associated with GTP and adenylate cyclase. The hormone-receptor complex leads to the cleavage of the βγ dimer from α GTP. The α subunit of GTP activates adenylate cyclase, which catalyzes the formation of cyclic AMP (c. AMP). c. AMP activates protein kinase A (PKA), which phosphorylates enzymes that change the rate of metabolic processes. Protein kinases are classified as A, B, C, etc.
 Adrenaline and glucagon, through the adenylate cyclase hormonal signal transmission system, activate hormone-dependent adipocyte TAG lipase. Occurs when the body is stressed (fasting, prolonged muscular work, cooling). Insulin blocks this process. Protein kinase A phosphorylates TAG lipase and activates it. TAG lipase cleaves fatty acids from triacylglycerols to form glycerol. Fatty acid oxidize and provide the body with energy.
Adrenaline and glucagon, through the adenylate cyclase hormonal signal transmission system, activate hormone-dependent adipocyte TAG lipase. Occurs when the body is stressed (fasting, prolonged muscular work, cooling). Insulin blocks this process. Protein kinase A phosphorylates TAG lipase and activates it. TAG lipase cleaves fatty acids from triacylglycerols to form glycerol. Fatty acid oxidize and provide the body with energy.
 Signal transmission from adrenergic receptors. AC – adenylate cyclase, Pk. A – protein kinase A, Pk. C – protein kinase C, Fl. C – phospholipase C, Fl. A 2 – phospholipase A 2, Fl. D – phospholipase D, PC – phosphatidylcholine, PL – phospholipids, FA – phosphatidic acid, Ach. K – arachidonic acid, PIP 2 – phosphatidylinositol biphosphate, IP 3 – inositol triphosphate, DAG – diacylglycerol, Pg – prostaglandins, LT – leukotrienes.
Signal transmission from adrenergic receptors. AC – adenylate cyclase, Pk. A – protein kinase A, Pk. C – protein kinase C, Fl. C – phospholipase C, Fl. A 2 – phospholipase A 2, Fl. D – phospholipase D, PC – phosphatidylcholine, PL – phospholipids, FA – phosphatidic acid, Ach. K – arachidonic acid, PIP 2 – phosphatidylinositol biphosphate, IP 3 – inositol triphosphate, DAG – diacylglycerol, Pg – prostaglandins, LT – leukotrienes.
 Adrenergic receptors of all types realize their action through Gs proteins. The α-subunits of this protein activate adenylate cyclase, which ensures the synthesis of c in the cell. AMP from ATP and activation of c. AMP-dependent protein kinase A. The ββ γ-subunit of the Gs protein activates L-type Ca 2+ channels and maxi-K+ channels. Under the influence of c. AMP-dependent protein kinase A phosphorylates myosin light chain kinase and it becomes inactive, unable to phosphorylate myosin light chains. The process of phosphorylation of light chains stops and the smooth muscle cell relaxes.
Adrenergic receptors of all types realize their action through Gs proteins. The α-subunits of this protein activate adenylate cyclase, which ensures the synthesis of c in the cell. AMP from ATP and activation of c. AMP-dependent protein kinase A. The ββ γ-subunit of the Gs protein activates L-type Ca 2+ channels and maxi-K+ channels. Under the influence of c. AMP-dependent protein kinase A phosphorylates myosin light chain kinase and it becomes inactive, unable to phosphorylate myosin light chains. The process of phosphorylation of light chains stops and the smooth muscle cell relaxes.
 American scientists Robert Lefkowitz and Brian Kobilka were awarded Nobel Prize in 2012 for understanding the mechanisms of interaction of adrenaline receptors with G-proteins. Interaction of the beta-2 receptor (indicated in blue) with G-proteins (indicated in green). G protein-coupled receptors are very beautiful if we consider the architectural molecular assemblies of the cell as masterpieces of nature. They are called “semi-spiral” because they are helically packed in the cell membrane in the manner of a Christmas tree serpentine and “pierce” it seven times, exposing a “tail” to the surface that is capable of receiving a signal and transmitting conformational changes to the entire molecule.
American scientists Robert Lefkowitz and Brian Kobilka were awarded Nobel Prize in 2012 for understanding the mechanisms of interaction of adrenaline receptors with G-proteins. Interaction of the beta-2 receptor (indicated in blue) with G-proteins (indicated in green). G protein-coupled receptors are very beautiful if we consider the architectural molecular assemblies of the cell as masterpieces of nature. They are called “semi-spiral” because they are helically packed in the cell membrane in the manner of a Christmas tree serpentine and “pierce” it seven times, exposing a “tail” to the surface that is capable of receiving a signal and transmitting conformational changes to the entire molecule.
 G proteins are a family of proteins that belong to GTPases and function as intermediaries in intracellular signaling cascades. G proteins are so named because in their signaling mechanism they use the replacement of GDP ( Blue colour) to GTF ( green color) as a molecular functional “switch” for regulating cellular processes.
G proteins are a family of proteins that belong to GTPases and function as intermediaries in intracellular signaling cascades. G proteins are so named because in their signaling mechanism they use the replacement of GDP ( Blue colour) to GTF ( green color) as a molecular functional “switch” for regulating cellular processes.
 G proteins are divided into two main groups - heterotrimeric (“large”) and “small”. Heterotrimeric G proteins are proteins with a quaternary structure, consisting of three subunits: alpha (α), beta (β) and gamma (γ). Small G proteins are proteins from a single polypeptide chain, they have molecular weight 20-25 k. Yes, and they belong to the Ras superfamily of small GTPases. Their single polypeptide chain is homologous to the α subunit of heterotrimeric G proteins. Both groups of G proteins are involved in intracellular signaling.
G proteins are divided into two main groups - heterotrimeric (“large”) and “small”. Heterotrimeric G proteins are proteins with a quaternary structure, consisting of three subunits: alpha (α), beta (β) and gamma (γ). Small G proteins are proteins from a single polypeptide chain, they have molecular weight 20-25 k. Yes, and they belong to the Ras superfamily of small GTPases. Their single polypeptide chain is homologous to the α subunit of heterotrimeric G proteins. Both groups of G proteins are involved in intracellular signaling.
 Cyclic adenosine monophosphate (cyclic AMP, c. AMP, c. AMP) is an ATP derivative that acts as a secondary messenger in the body, used for the intracellular distribution of signals of certain hormones (for example, glucagon or adrenaline) that cannot pass through the cell membrane. .
Cyclic adenosine monophosphate (cyclic AMP, c. AMP, c. AMP) is an ATP derivative that acts as a secondary messenger in the body, used for the intracellular distribution of signals of certain hormones (for example, glucagon or adrenaline) that cannot pass through the cell membrane. .
 Each of the hormonal signal transmission systems corresponds to a specific class of protein kinases. The activity of type A protein kinases is regulated by c. AMP, protein kinase G - c. GMF. Ca 2+ - calmodulin-dependent protein kinases are controlled by the concentration of CA 2+. Type C protein kinases are regulated by DAG. An increase in the level of any second messenger leads to the activation of a certain class of protein kinases. Sometimes a membrane receptor subunit may have enzyme activity. For example: insulin receptor tyrosine protein kinase, whose activity is regulated by the hormone.
Each of the hormonal signal transmission systems corresponds to a specific class of protein kinases. The activity of type A protein kinases is regulated by c. AMP, protein kinase G - c. GMF. Ca 2+ - calmodulin-dependent protein kinases are controlled by the concentration of CA 2+. Type C protein kinases are regulated by DAG. An increase in the level of any second messenger leads to the activation of a certain class of protein kinases. Sometimes a membrane receptor subunit may have enzyme activity. For example: insulin receptor tyrosine protein kinase, whose activity is regulated by the hormone.
 The action of insulin on target cells begins after it binds to membrane receptors, and the intracellular domain of the receptor has tyrosine kinase activity. Tyrosine kinase triggers the phosphorylation of intracellular proteins. The resulting autophosphorylation of the receptor leads to an increase in the primary signal. The insulin receptor complex can cause activation of phospholipase C, formation of second messengers inositol triphosphate and diacylglycerol, activation of protein kinase C, inhibition of c. AMF. The involvement of several second messenger systems explains the diversity and differences in the effects of insulin in different tissues.
The action of insulin on target cells begins after it binds to membrane receptors, and the intracellular domain of the receptor has tyrosine kinase activity. Tyrosine kinase triggers the phosphorylation of intracellular proteins. The resulting autophosphorylation of the receptor leads to an increase in the primary signal. The insulin receptor complex can cause activation of phospholipase C, formation of second messengers inositol triphosphate and diacylglycerol, activation of protein kinase C, inhibition of c. AMF. The involvement of several second messenger systems explains the diversity and differences in the effects of insulin in different tissues.
 Another system is the guanylate cyclase messenger system. The cytoplasmic domain of the receptor has guanylate cyclase (heme-containing enzyme) activity. Molecules c. GTP can activate ion channels or protein kinase GG, which phosphorylate enzymes. c. GMP controls water exchange and ion transport in the kidneys and intestines, and serves as a relaxation signal in the heart muscle.
Another system is the guanylate cyclase messenger system. The cytoplasmic domain of the receptor has guanylate cyclase (heme-containing enzyme) activity. Molecules c. GTP can activate ion channels or protein kinase GG, which phosphorylate enzymes. c. GMP controls water exchange and ion transport in the kidneys and intestines, and serves as a relaxation signal in the heart muscle.

 Inositol phosphate system. The binding of a hormone to a receptor causes a change in the conformation of the receptor. Dissociation of the G-G protein occurs and GDP is replaced by GTP. The separated α-subunit associated with the GTP molecule acquires an affinity for phospholipase C. Under the action of phospholipase-C, the membrane lipid phosphatidylinositol-4, 5-bisphosphate (PIP 2) is hydrolyzed and inositol-1, 4, 5-triphosphate (IP 3) is formed ) and diacylglycerol (DAG). DAG is involved in the activation of the enzyme protein kinase C (PKC). Inositol-1, 4, 5-triphosphate (IP 3) binds to specific centers of the Ca 2+ channel of the ER membrane, this leads to a change in protein conformation and opening of the channel - Ca 2+ enters the cytosol. In the absence of IF in the cytosol, channel 3 is closed.
Inositol phosphate system. The binding of a hormone to a receptor causes a change in the conformation of the receptor. Dissociation of the G-G protein occurs and GDP is replaced by GTP. The separated α-subunit associated with the GTP molecule acquires an affinity for phospholipase C. Under the action of phospholipase-C, the membrane lipid phosphatidylinositol-4, 5-bisphosphate (PIP 2) is hydrolyzed and inositol-1, 4, 5-triphosphate (IP 3) is formed ) and diacylglycerol (DAG). DAG is involved in the activation of the enzyme protein kinase C (PKC). Inositol-1, 4, 5-triphosphate (IP 3) binds to specific centers of the Ca 2+ channel of the ER membrane, this leads to a change in protein conformation and opening of the channel - Ca 2+ enters the cytosol. In the absence of IF in the cytosol, channel 3 is closed.

Based on their mechanism of action, hormones are divided into two main types. The first is protein and peptide hormones, catecholamines and hormonoids. Their molecule, approaching the target cell, connects with the protein receptor molecules of the outer plasma membrane, then, with the help of mediators (c AMP, c GMP, prostaglandins, Ca2+), it influences the enzyme systems of the target cell and the metabolism in it. The second type of hormones includes steroid hormones and some thyroid hormones. Their molecule easily penetrates deep into the target cell through the pores of the membrane; interacts with molecules of glycoprotein receptors localized in the cytosol, mitochondria on the nuclear membrane, affecting the entire cellular metabolism, and primarily transcription processes.
Mechanisms of action of hormones on target cells.
Depending on the structure of the hormone, there are two types of interaction. If the hormone molecule is lipophilic (for example, steroid hormones), then it can penetrate the lipid layer outer membrane target cells. If the molecule is large or polar, then its penetration into the cell is impossible. Therefore, for lipophilic hormones, the receptors are located inside the target cells, and for hydrophilic hormones, the receptors are located in the outer membrane.
To obtain a cellular response to a hormonal signal in the case of hydrophilic molecules, it acts intracellular mechanism signal transmission. This occurs with the participation of substances called second messengers. Hormone molecules are very diverse in shape, but “second messengers” are not.
The reliability of signal transmission is ensured by the very high affinity of the hormone for its receptor protein.
What are the intermediaries that are involved in the intracellular transmission of humoral signals?
These are cyclic nucleotides (cAMP and cGMP), inositol triphosphate, calcium-binding protein - calmodulin, calcium ions, enzymes involved in the synthesis of cyclic nucleotides, as well as protein kinases - protein phosphorylation enzymes. All these substances are involved in the regulation of the activity of individual enzyme systems in target cells.
Let us examine in more detail the mechanisms of action of hormones and intracellular mediators.
There are two main ways of transmitting a signal to target cells from signaling molecules with a membrane mechanism of action:
adenylate cyclase (or guanylate cyclase) systems;
phosphoinositide mechanism.
Before finding out the role of the cyclase system in the mechanism of action of hormones, let's consider the definition of this system. The cyclase system is a system consisting of adenosine cyclophosphate, adenylate cyclase and phosphodiesterase contained in the cell, which regulates the permeability of cell membranes, is involved in the regulation of many metabolic processes of a living cell, and mediates the action of some hormones. That is, the role of the cyclase system is that they are second intermediaries in the mechanism of action of hormones.
The adenylate cyclase - cAMP system. The membrane enzyme adenylate cyclase can be found in two forms - activated and non-activated. Activation of adenylate cyclase occurs under the influence of a hormone-receptor complex, the formation of which leads to the binding of guanyl nucleotide (GTP) to a special regulatory stimulating protein (GS protein), after which the GS protein causes the addition of magnesium to adenylate cyclase and its activation. This is how the hormones activating adenylate cyclase act: glucagon, thyrotropin, parathyrin, vasopressin, gonadotropin, etc. Some hormones, on the contrary, suppress adenylate cyclase (somatostatin, angiotensin-P, etc.).
Under the influence of adenylate cyclase, cAMP is synthesized from ATP, which causes activation of protein kinases in the cell cytoplasm, which ensure the phosphorylation of numerous intracellular proteins. This changes the permeability of membranes, i.e. causes metabolic and, accordingly, functional changes typical for the hormone. The intracellular effects of cAMP are also manifested in their influence on the processes of proliferation, differentiation, and the availability of membrane receptor proteins to hormone molecules.
"Guanylate cyclase - cGMP" system. Activation of membrane guanylate cyclase occurs not under the direct influence of the hormone-receptor complex, but indirectly through ionized calcium and oxidative membrane systems. This is how atrial natriuretic hormone - atriopeptide, tissue hormone - realizes its effects vascular wall. In most tissues, the biochemical and physiological effects of cAMP and cGMP are opposite. Examples include stimulation of cardiac contractions under the influence of cAMP and inhibition of them by cGMP, stimulation of contractions of intestinal smooth muscles by cGMP and inhibition of cAMP.
In addition to the adenylate cyclase or guanylate cyclase systems, there is also a mechanism for transmitting information within the target cell with the participation of calcium ions and inositol triphosphate.
Inositol triphosphate is a substance that is a derivative of a complex lipid - inositol phosphatide. It is formed as a result of the action of a special enzyme - phospholipase "C", which is activated as a result of conformational changes in the intracellular domain of the membrane receptor protein.
This enzyme hydrolyzes the phosphoester bond in the phosphatidyl-inositol 4,5-bisphosphate molecule to form diacylglycerol and inositol triphosphate.
It is known that the formation of diacylglycerol and inositol triphosphate leads to an increase in the concentration of ionized calcium inside the cell. This leads to the activation of many calcium-dependent proteins inside the cell, including the activation of various protein kinases. And here, as with the activation of the adenylate cyclase system, one of the stages of signal transmission inside the cell is protein phosphorylation, which leads to a physiological response of the cell to the action of the hormone.
A special calcium-binding protein, calmodulin, takes part in the phosphoinositide signaling mechanism in the target cell. This is a low molecular weight protein (17 kDa), 30% consisting of negatively charged amino acids (Glu, Asp) and therefore capable of actively binding Ca+2. One calmodulin molecule has 4 calcium-binding sites. After interaction with Ca+2, conformational changes occur in the calmodulin molecule and the “Ca+2-calmodulin” complex becomes capable of regulating the activity (allosterically inhibiting or activating) many enzymes - adenylate cyclase, phosphodiesterase, Ca+2,Mg+2-ATPase and various protein kinases.
In different cells, when the Ca+2-calmodulin complex acts on isoenzymes of the same enzyme (for example, adenylate cyclase different types) in some cases activation is observed, and in others inhibition of the cAMP formation reaction is observed. These different effects occur because the allosteric centers of the isoenzymes may include different amino acid radicals and their response to the action of the Ca+2-calmodulin complex will be different.
Thus, the role of “second messengers” for transmitting signals from hormones in target cells can be:
cyclic nucleotides (c-AMP and c-GMP);
complex "Ca-calmodulin";
diacylglycerol;
inositol triphosphate.
The mechanisms for transmitting information from hormones inside target cells using the listed intermediaries have common features:
one of the stages of signal transmission is protein phosphorylation;
cessation of activation occurs as a result of special mechanisms initiated by the process participants themselves - there are negative feedback mechanisms.
Hormones are the main humoral regulators physiological functions organism, and their properties, biosynthesis processes and mechanisms of action are now well known. Hormones are highly specific substances in relation to target cells and have very high biological activity.
As a result of interaction with receptors in target cells, specific hormonal effects can be mediated by three main mechanisms, namely:
1) direct influence on membrane processes;
2) systems of intracellular “second messengers”;
3) action on the cell nucleus.
However, it must be borne in mind that one hormone can act on several by different mechanisms. A number of hormones can be distinguished fast(metabolic) and slow(growth) impact. For example, insulin causes in muscles quick changes in the transport and metabolism of sugars and amino acids and long-term, slow changes in the synthesis and metabolism of proteins.
For fast effects, the mechanism is rather the activation of the enzymatic apparatus of cell membranes; slow effects require the interaction of the nuclear genome.
2.2.1. Direct membrane effects
Hormones can have a direct effect on the plasma membranes of cells:
a) change the permeability of membranes to ions or the transport of certain compounds (for example, the effect of insulin on the transport of glucose and amino acids through membranes);
b) change the structure of the membrane (for example, open pores);
c) change the activity of carriers (for example, by changing their conformation and affinity for the transported substances);
d) stimulate the formation of specific “pores” or “channels” in the membrane;
e) activate specific membrane “pumps”, for example, the iodide pump in thyroid cells.
2.2.2. Activation of intracellular second messengers
The biological effect of hormones interacting with receptors localized on the plasma membrane is carried out using special substances - secondary transmitters or messengers. It is currently known that the role of messengers can be played by at least, the following substances: cyclic adenosine-3′,5′-monophosphate (cAMP) and cyclic guanosine-3′,5′-monophosphate (cGMP), inositol triphosphate, diacylglycerol, calcium ions, eicosanoids and some other factors of unknown nature.
Work of c-AMP as a messenger.
C-AMP is formed in the cell under the influence of the enzyme adenylate cyclase from ATP molecules. Therefore, the main effect of the hormone should be aimed at changing the activity of adenylate cyclase. Adenylate cyclase consists of three components: a receptor, a regulatory protein and a catalytic subunit, which are separated from each other in the unstimulated state. The receptor is located on outside membranes. The regulatory unit is represented by a g-protein and is located on inner surface plasma membrane. In the absence of the hormone, it is bound to guanosine diphosphate (GDP). When a hormone acts on the receptor part, the subunit binds to guanosite triphosphate and is activated. The role of the hormone is to replace the g-protein–GDP complex with the g-protein–GTP complex. As a result, the content of c-AMP increases. The resulting c-AMP activates protein kinases. Each protein kinase molecule consists of two regulatory and two catalytic subunits. CAMP causes dissociation of protein kinase subunits, free catalytic subunits are able to phosphorylate specific protein substrates, thereby realizing the intracellular effects of hormones. (Table 4).
Table 3
Hormones whose effects on tissue are mediated by cAMP
So: hormone + receptor ® activation of adenylate cyclase ® activation of protein kinase ® protein phosphorylation → intracellular effects of the hormone.
Cyclic GMP (cGMP)
CAMP is formed due to the activation of the membrane catalytic unit, guanyl cyclase. Unlike adenylate cyclase, guanyl cyclase simultaneously functions as a receptor and a catalytic unit. Examples of hormones that directly interact with membrane guanyl cyclase and mediate their effects through cGMP are atrial natriuretic peptides and nitric oxide.
Phosphoinositides
When a hormone binds to a membrane receptor, a system of second messengers formed from membrane phospholipids can be activated. The receptor, in such cases, is in a complex with the G-protein and when the receptor interacts with the hormone, the membrane enzyme (phospholipase C) is activated. Acting on membrane phospholipids, namely phosphatidylinositol-4,5-biphosphate (PIP 2), this enzyme leads to the formation inositol triphosphate(IF 3) and diacylglycerol(DAG) (Fig. 5). These compounds then act as second messengers, influencing intracellular calcium levels.
Rice. 5. An example of activation of membrane phospholipase by a hormone.
The binding of a hormone to its membrane receptor can lead to the activation of membrane phospholipase C (PLS), under the action of which inositol triphosphate (IP 3) and diacylglycerol (DAG) are formed from phosphaimdylinositol diphosphate (PIP 2). IP3 enhances the movement of calcium ions from intracellular stores into the cytoplasm, and DAG activates protein kinase C (PKC).
Calcium, which is mobilized from reserves sensitive to IF3, stimulates its release from other (insensitive to IF3) intracellular reserves, as a result of which a “wave” of this ion quickly spreads throughout the cytoplasm. An increase in the level of calcium in the cytoplasm means the appearance of another intracellular mediator, since calcium ions have numerous effects on metabolic processes.
IP 3 produces various other phosphorylated forms of inositol, most of which are inactive, although some may enhance the intracellular effects of IP 3.
FIF 2 also produces diacylglycerol(DAG), which activates the membrane enzyme protein kinase C (PKC). This enzyme phosphorylates intracellular proteins, which can then influence various metabolic processes (both in the cytoplasm and in the nucleus), causing the manifestation of hormonal effects. Activation of PKC under the influence of DAG can also enhance the work of the calcium pump in the cell membrane, which ensures restoration of the initial level of calcium in the cytoplasm.
The messenger is calcium ions.
The process of activation of protein kinases is also associated with the interaction of calcium ions with the cell regulatory protein – calmodulin. Typically, calmodulin is in an inactive state and therefore is not able to exert its regulatory effect on enzymes. In the presence of calcium, calmodulin is activated, resulting in the activation of protein kinases, which subsequently leads to phosphorylation of proteins.
The role of the hormone in in this case consists in changing the permeability of the cell membrane for calcium ions or due to the release of free calcium ions from intracellular
depot (Fig. 5).
Increased level intracellular calcium is eliminated by stimulating the calcium pump, which “pumps” free calcium into the intercellular fluid, reducing its level in the cell, as a result of which calmodulin becomes inactive and the state of functional rest is restored in the cell.
So: hormone + receptor ® increased calcium level in the cell ® activation of calmodulin ® activation of protein kinase ® phosphorylation of the regulatory protein ® physiological effect.
Other messengers.
Mediators hormonal action may also be arachidonic acid. The interaction of the hormone with the receptor promotes the destruction of membrane phospholipids and increased formation of arachidonic acid and prostaglandins, which mediate the hormonal effect.
The synthesis of prostaglandins passes through the formation of unstable intermediate products - endoperoxides, which serve as precursors to other biological substances. active compounds– thromboxanes. Other active molecules, prostacyclins, are also formed from endoperoxides.
Arachidonic acid is also a precursor to another group of active compounds - leukotrienes, which are synthesized in blood leukocytes. Unlike prostaglandins and thromboxanes, which act primarily as intracellular messengers, leukotrienes and prostacyclin are released from cells into the blood and can be considered hormones.
2.2.3. Action on the cell nucleus
For most genes, regulated by hormones, characterized by the presence of nucleotide sequences that act as hormone-binding elements. As a result of the binding of the hormone to the target DNA, the transcription process changes and, ultimately, the desired protein molecule is synthesized. Transcriptional repression can also occur.
There are two stages in the protein synthesis process that can be influenced by hormones:
Transcription of code from DNA to RNA;
Translation of the mRNA code during protein synthesis on ribosomes.
Thyroid and steroid hormones cortisol and estrogens stimulate protein synthesis at the transcription stage. Other hormones that stimulate protein synthesis in the cell influence protein synthesis at the translation stage.

Orange juice. The symbolic meaning of orange juice in dream books is pleasure and temptation. Quite often we...

We most often overcome all kinds of difficulties encountered along the path of life. Of course, for this we make...

This year your patron Neptune will be in your constellation and this is a good sign, because you will feel...

1993 who? 1993 is the year of which animal? — According to the Chinese horoscope, 1873, 1933, 1993 belonged to the years of the Black Water...

Wave-particle duality of light means that light simultaneously has the properties of continuous electromagnetic...

The role of biology is enormous in our world. Although it is not a priority subject, most students and parents...
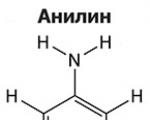
Amines are organic derivatives of ammonia containing an NH 2 amino group and an organic radical. In general...
How to answer questions in Part BThe second part of the social studies work consists of 7 tasks with a short answer....

Form TORG-15 is drawn up in the case when during transportation, movement between and within the warehouse, when...

Nutritionists say that for good health and a slim figure, you must include snacks in your...

Delicious pickled carrots for the winter can be prepared in a variety of containers, it can be a wooden...

When I have a few minutes left to prepare breakfast, the simplest and fastest recipes are used....

An elegant table, a decorated Christmas tree, tangerine spirit spilled throughout all the rooms, soon the most magical holiday -...

Each of us has repeatedly faced financial difficulties and difficult periods in life when Fortune...

We most often overcome all kinds of difficulties encountered along the path of life. Of course, for this we...

This year your patron Neptune will be in your constellation and this is a good sign, because you will...