Kas galima ir ko negalima per Gimimo pasninką?
2018 metais gimimo pasninkas prasidės lapkričio 28 d. Šiuo laikotarpiu stačiatikiai ruošiasi švęsti Kalėdas...
C ir G grupės. Proteinas G panašus į baltymą A, tačiau skiriasi specifiškumu. G baltymo molekulinė masė yra 58 kDa (C40 baltymo atveju) arba 65 kDa (G148 baltymo atveju). Baltymas G jungiasi prie antikūnų α regiono, todėl yra plačiai naudojamas imunoglobulinų valymui. G baltymų molekulės taip pat jungiasi su albuminu.
Yra ir kitų bakterijų baltymų, kurie suriša imunoglobulinus – baltymas A, baltymas A/G ir baltymas L. Šie baltymai naudojami imunoglobulinų gryninimui, imobilizavimui ir išskyrimui. Šie antikūnus surišantys baltymai turi skirtingus imunoglobulinų prisijungimo profilius.
Wikimedia fondas. 2010 m.
- (angl. protein A) – 40-60 kDa molekulinės masės baltymas, išskirtas iš auksinio stafilokoko (Staphylococcus aureus) ląstelės sienelės paviršiaus. Baltymas A naudojamas biocheminiuose tyrimuose, nes gerai suriša daugelį... ... Vikipedijos
Gawk į arabų voveres.. Rusų sinonimų ir panašių posakių žodynas. pagal. red. N. Abramova, M.: Rusų žodynai, 1999. baltymų globulinas, histonas, proteinoidas, baltymas, baltyminis kūnas, protaminas, proteidas Rusų sinonimų žodynas ... Sinonimų žodynas
Ušakovo aiškinamasis žodynas
1. BALTYMAS1, voverė, patinas. (biol. chem.). Svarbiausias gyvūnų ir augalų kūno komponentas; tas pats kaip albuminas ir baltymai. 2. BALTYMAS2, voverė, patinas. 1. Išgaubtas nepermatomas akies apvalkalas. || tik daugiskaita Akys (paprastos). Išsukite baltymus. Baltymai...... Ušakovo aiškinamasis žodynas
1. BALTYMAS1, voverė, patinas. (biol. chem.). Svarbiausias gyvūnų ir augalų kūno komponentas; tas pats kaip albuminas ir baltymai. 2. BALTYMAS2, voverė, patinas. 1. Išgaubtas nepermatomas akies apvalkalas. || tik daugiskaita Akys (paprastos). Išsukite baltymus. Baltymai...... Ušakovo aiškinamasis žodynas
baltymas C- Baltymai, serino proteazė, sintetinami kepenų ląstelių; antikoaguliantas, yra kraujo krešėjimo faktorių Va ir VIIIa inhibitorius; heterozigotų dažnis dėl B.C trūkumo žmonių populiacijose siekia 1/200; ši anomalija yra susijusi su padidėjusiu... Techninis vertėjo vadovas
BALTYMAS, organinis JUNGINIS, turintis daug aminorūgščių, sujungtų kovalentinėmis peptidinėmis jungtimis. Baltymų molekulės susideda iš polipeptidinių grandinių. Gyvose ląstelėse yra apie 20 skirtingų aminorūgščių. Dėl to, kad kiekviename...... Mokslinis ir techninis enciklopedinis žodynas
BALTYMAS, lka, vyras. Didelės molekulinės masės organinė medžiaga, užtikrinanti gyvybines gyvūnų ir augalų organizmų funkcijas. | adj. baltymai, oi, oi. Baltyminiai pašarai (didelis baltymų kiekis). II. BALTYMAS, lka, vyras. 1. Permatoma kiaušinio dalis... Ožegovo aiškinamasis žodynas
Riebalų rūgštis surišantis baltymas (FABP) yra riebalų rūgščių ir kitų lipofilinių medžiagų, tokių kaip eikozanoidai ir retinoidai, pernešėjų šeima. Manoma, kad šie... ... Vikipedija
1. BALTYMAS, lka; m. 1. Skaidrus skystis, supantis paukščio kiaušinio trynį. / Apie šią vištienos kiaušinio dalį kaip maistą. Gerti žalią b. Išplakti baltymai. ◁ Baltymai, oi, oi. B. grietinėlė (iš kiaušinių baltymų). 2. BALTYMAI žr. 1. Baltymai. 3. BALTYMAI žr. 2. Baltymai. 4.…… enciklopedinis žodynas
(Anglų) guanino nukleotidus surišantys baltymai, Guanilo nukleotidus surišantys baltymai yra baltymų šeima, dalyvaujanti eukariotinių ląstelių signalizacijoje. G baltymai atlieka savotiškų jungiklių vaidmenį: jie gali persijungti iš neaktyvios būsenos į aktyvią ir atvirkščiai, atitinkamai įjungdami arba išjungdami tam tikro signalo perdavimą ląstelės viduje. Šie baltymai gavo savo pavadinimą dėl jų gebėjimo surišti guanilo nukleotidus. G uanino nukleotidas): komplekse su guanozino difosfatu (BVP) jie yra neaktyvūs, o komplekse su guanozino trifosfatu (GTP) – aktyvūs.
Terminas „G baltymai“ dažniau vartojamas kalbant apie heterotrimerinius (stambius) GTP surišančius baltymus, susidedančius iš trijų subvienetų α, β ir γ; Yra dar viena GTP surišančių baltymų klasė – monomerai, kurie kartais vadinami mažais G-baltymais (Ras mažųjų GTPazių supernamu), jie yra homologiški didelių α-subvienetams.
Heterotrimeriniai G baltymai dalyvauja perduodant signalus iš su G baltymu susietų receptorių. G-baltymu susieti receptoriai (GPCR)- didžiausia ląstelių receptorių klasė (pavyzdžiui, in Caenorhabditis elegantiškas jų genai užima 5 % viso genomo). Stuburiniams gyvūnams jie yra atsakingi už daugelio hormonų ir kitų signalinių molekulių suvokimą, taip pat už cheminius pojūčius (uoslę ir skonį) ir fotorecepciją (regėjimą). Svarbu tai, kad maždaug pusė žinomų farmacinių vaistų veikia per G baltymus susietus receptorius: tai gerai žinomi vaistai, tokie kaip antihistamininiai vaistai Claritin (loratadinas) ir antidepresantas Prozac (fluoksetinas), taip pat psichotropinės medžiagos, ypač heroinas, kokainas ir tetrahidrokanabinolis (veiklioji marihuanos medžiaga).
Heterotrimerinius G baltymus atrado Alfredas Gilmanas ir Martinas Rodbellas, už kuriuos 1994 m. jie gavo Nobelio fiziologijos ar medicinos premiją.
Hetrotrimeriniai G baltymai susideda iš trijų subvienetų: α, β ir γ. α subvienete yra GTP surišimo ir hidrolizės domenas, kuris yra identiškas visoje GTPazės superhomelande. β-subvienete yra 7 β-struktūros, išdėstytos kaip sraigto mentės. γ-subvienetas glaudžiai sąveikauja su β-subvienetu; kartu jie sudaro vieną funkcinę struktūrą, kuri gali atsiskirti tik baltymų hidrolizės atveju. Visas G baltymas yra pritvirtintas prie membranos dviem lipidais, iš kurių vienas yra kovalentiškai prijungtas prie α-subvieneto N-galo, kitas - prie γ-subvieneto C-galo.
G baltymu sujungti receptoriai G-baltymu susieti receptoriai, GPCR)- didžiausia eukariotų ląstelių receptorių šeima, užtikrinanti hormonų, neurotransmiterių, vietinių reguliatorių suvokimą, taip pat stuburiniams gyvūnams regėjimą, kvapą ir skonio pojūtį. Žmogaus genome randama apie 700 GPCR genų, o pelės vien uoslės receptorių yra daugiau nei 1000.
Signalinės molekulės, veikiančios kaip su G baltymu susietų receptorių ligandai, gali būti labai skirtingos cheminės prigimties: baltymai, smulkūs peptidai, lipidai, aminorūgščių dariniai ir panašiai. Be to, kai kurie šios klasės receptorių nariai, ypač rodopsinas, gali suvokti šviesos fotonus. Kartais vienai signalizacijos molekulei egzistuoja keli skirtingi GPCR, jie yra išreikšti skirtinguose ląstelių tipuose ir sukelia skirtingus signalizacijos kelius. Pavyzdžiui, žmogaus organizme yra mažiausiai 9 skirtingi adrenalino receptoriai ir mažiausiai 14 neurotransmiterio serotonino.
Visi su G baltymu sujungti receptoriai turi panašią struktūrą: jie susideda iš vienos polipeptidinės grandinės, kuri 7 kartus kerta lipidų dvisluoksnį. Kiekvienas transmembraninis domenas yra pavaizduotas α-spirale, kurią sudaro 20-30 nepolinių aminorūgščių. Šie domenai yra tarpusavyje sujungti įvairaus dydžio kilpomis, esančiomis abiejose plazminės membranos pusėse. GPCR daugiausia yra glikoproteinai, kurių angliavandenių liekanos yra glikoproteino pusėje. Šių receptorių viduląsteliniuose domenuose yra sąveikos su G baltymais vietos.
G baltymai atlieka ląstelių receptorių sujungimo su tam tikromis efektorinėmis molekulėmis, tokiomis kaip fermentai ar jonų kanalai, vaidmenį, o veikia kaip molekuliniai jungikliai. Neaktyvioje būsenoje G baltymuose yra BVP, susieto su α subvienetu.
Signalo perdavimas prasideda, kai atitinkamas ligandas veikia ląstelės receptorių, ko pasekoje receptorius suaktyvėja ir keičia konformaciją. Suaktyvintas receptorius veikia G baltymą (kuris arba visam laikui susikompleksuoja su juo, arba susijungia po aktyvacijos), todėl α subvieneto struktūra pasikeičia taip, kad jis atpalaiduoja surištą BVP molekulę. Šios molekulės vietą greitai užima GTP, dėl to suaktyvėja G-baltymas ir pasikeičia jo struktūra: α-subvienetas praranda afinitetą βγ kompleksui ir jis suyra. Esant tokiai aktyvuotai būsenai, tiek su GTP surištas α-subvienetas, tiek βγ-kompleksas gali vykdyti signalo perdavimą: aktyvuoti tam tikrus fermentus arba paveikti jonų kanalų būseną. α-subvienetas yra GTPazė, ir kai tik jis hidrolizuoja prijungtą GTP į BVP, jis iš karto inaktyvuojamas ir atkuriama G-baltymo trimerio struktūra. Tokiu būdu garsas išjungiamas. Inaktyvuotas G baltymas gali sąveikauti su vėlesne receptoriaus molekule ir vėl įsijungti.
Tam tikro signalo perdavimo per G baltymą efektyvumas priklauso nuo aktyvių, su GTP surištų ir neaktyvių, su BVP susijusių formų koncentracijos santykio. Ir šis santykis, savo ruožtu, priklauso nuo dviejų būsenų: BVP disociacijos konstantos ir GTP hidrolizės greičio konstantos:
Kur
Šį ryšį patvirtina GTP perteklius terpėje, taip pat greitas, beveik momentinis jo prisijungimas prie „tuščios“ G baltymo molekulės (ty nesusijusios su vienu guanilo nukleotidu). Tokiu atveju signalo perdavimo efektyvumą galima reguliuoti vienu iš šių būdų:
G baltymai gauna įvestį iš su jais susijusių receptorių, po kurių jie aktyvuoja vieną iš ląstelės signalizacijos kelių.
Ciklinis AMP (cAMP) yra įprastas antrasis pasiuntinys, valdantis daugelį procesų eukariotinėse ląstelėse. cAMP sintetina su ATP dideliu transmembraniniu fermentu adenilato ciklaze ir skaido cAMP fosfodiesterazė. Daugelis signalizuojančių molekulių veikia ląstelę, padidindamos arba sumažindamos cAMP koncentraciją aktyvindamos arba slopindamos adenilato ciklazę. cAMP atlieka savo funkciją kaip antrasis pasiuntinys, aktyvuodamas nuo cAMP priklausomą proteinkinazę (protein kinase A, PKA), kuri savo ruožtu fosforilina daugybę baltymų ląstelėje ties serino ir treonino liekanomis, juos aktyvuodama arba deaktyvuodama.
Yra dviejų tipų G baltymai, kurie veikia aktyvias adenilato ciklazes: G s (angl. Stimuliuojantis)- stimuliuoja, aktyvina ir didina cAMP ir G i koncentraciją (angl. Slopinantis)- slopina, slopina adenilato ciklazę, bet taip pat veikia tiesiogiai veikdamas jonų kanalus. Reakcijų, kurias sukelia nuo Gs priklausomas cAMP koncentracijos padidėjimas, pavyzdžiai:
G baltymai, turintys įtakos nuo cAMP priklausomam ląstelių signalizavimui, yra bakterijų toksinų taikiniai:
Šie du toksinai naudojami biologiniuose tyrimuose, siekiant nustatyti, kurį ląstelių atsaką tarpininkauja G s arba G i baltymas.
Daugelis su G baltymu susietų receptorių veikia aktyvuodami fosfolipazę C-β (PLC-β). Šis fermentas veikia inozitolio fosfolipidą: fosfatidilinozitolio 4,5 bisfosfatą (PI(4,5)P2 arba PIF 2), kurio nedideliais kiekiais yra plazmos membranos lipidinio dvisluoksnio vidiniame lapelyje. Receptoriai, aktyvuojantys šį signalizacijos kelią, paprastai yra sujungti su G q -bloku, kuris aktyvuoja fosfolipazę C panašiai kaip G s blokas - adenilato ciklazė. Aktyvuota fosfolipazė skaido fosfatidilinozitolio 4,5 bisfosfatą iki inozitolio 1,4,5 trifosfato (IP 3) ir diaciglicerolio (DAG). Šiame etape signalizacijos kelias išsišakoja:
Nuo G baltymo priklausomo fosfolipazės C-β aktyvavimo ląstelių reakcijų pavyzdžiai:
Daugelis G baltymų veikia atidarydami arba uždarydami jonų kanalus, taip pakeisdami elektrines plazmos membranos savybes.
Pavyzdžiui, širdies susitraukimų dažnis ir stiprumas, veikiant acetilcholinui, sumažėja dėl to, kad muskarininiai acetilcholino receptoriai po aktyvavimo sąveikauja su G i -subvienetu, kurio α-subvienetas slopina adenilato aktyvumą. ciklazė, o βγ kompleksas atveria kalio kanalus širdies ląstelių raumenų plazminėje membranoje, dėl ko sumažėja jų jaudrumas.
Kiti G baltymai jonų kanalų veiklą reguliuoja netiesiogiai: pavyzdžiui, regos ir uoslės receptoriai veikia per G baltymus, kurie veikia ciklinių nukleotidų sintezę, savo ruožtu uždarydami arba atidarydami jonų kanalus (jonų kanalus, valdomus cikliniais nukleotidais). Pavyzdžiui, visi uoslės receptoriai yra susiję su G olf baltymu, kuris aktyvina adenilato ciklazę; Sintetinamas cAMP atveria natrio kanalus, o tai lemia membranos depoliarizaciją ir nervinio impulso (receptoriaus potencialo), kuris perduodamas neuronams, generavimą.
Žmogaus tinklainės lazdelėse šviesai jautri molekulė yra rodopsinas. Šių ląstelių plazmos membranoje yra daug cGMP valdomų katijonų kanalų. Nesant šviesos stimuliacijos, strypų citoplazmoje yra didelis cGMP kiekis, kuris palaiko katijonų kanalus atvirus. Dėl to membrana periodiškai depoliarizuojasi ir vyksta sinapsinis impulsų perdavimas neuronams. Po aktyvavimo šviesa rodopsinas keičia konformaciją ir sąveikauja su G baltymo transducinu (Gt). Po to jo α-subvienetas aktyvuoja cGMP fosfodiesterazę, kuri suardo cGMP, dėl ko užsidaro katijonų kanalai ir sustoja sinapsinis perdavimas. Būtent iš šviesai jautrių ląstelių ateinančių impulsų dažnio sumažėjimą smegenys suvokia kaip šviesos pojūtį.
Visi heterotrimeriniai G baltymai yra suskirstyti į keturias pagrindines šeimas pagal α subvieneto aminorūgščių seką:
| Pagrindinės heterotrimerinių G baltymų šeimos, pagrįstos α subvieneto aminorūgščių seka | |||
|---|---|---|---|
| Šeima | Kai kurie nariai | Subvienetas, atsakingas už poveikį | Kai kurios funkcijos |
| aš | G s | α | Adenilato ciklazės aktyvinimas, kalcio kanalų atidarymas |
| G olf | α | Adenilato ciklazės aktyvinimas uoslės neuronuose | |
| II | G i | α | Adenilato ciklazės slopinimas |
| βγ | Kalio kanalų atidarymas | ||
| G o | βγ | Kalio kanalų atidarymas, kalcio kanalų uždarymas | |
| α ir βγ | Fosfolipazės C-β aktyvinimas | ||
| Gt (transducinas) | α | cGMP fosfodiesterazės aktyvinimas stuburinių fotoreceptoriuose | |
| III | Gq | α | Fosfolipazės C-β aktyvinimas |
| IV | G 12/13 | α | Rho šeimos monomerinių GTPazių, reguliuojančių aktino citoskeletą, aktyvinimas |
(C baltymu sujungti receptoriai, GPCR), perduoda signalą iš pirminių pasiuntinių į tarpląstelinius taikinius, naudojant GPCR-^-G-baltymo^ efektoriaus baltymų kaskadą. Pagrindiniai šių receptorių signalai yra daugybė molekulių, pavyzdžiui, mažos molekulinės masės hormonai ir neurotransmiteriai (tokie kaip adrenalinas, norepinefrinas, acetilcholinas, serotoninas, histaminas), opioidai, peptidai ir baltymų hormonai (adrenokortikotropinas, somaostatinas, vazopresinas, angiopresinas). , gonadotropinas , epidermio augimo faktorius), kai kurie neuropeptidai.
Toje pačioje serijoje yra daug cheminių signalų, kuriuos suvokia uoslės ir skonio jutimo ląstelės, ir šviesa, kurios receptorius yra regos arba fotoreceptorių ląstelių pigmentas rodopsinas.
Reikėtų atsižvelgti į tai, kad tas pats pirminis signalas gali inicijuoti signalo perdavimą per kelis (kartais daugiau nei 10) skirtingų GPCR, todėl jei išorinių signalų skaičius GPCR yra keliasdešimt, tai yra žinoma daugiau nei 200 pačių tokių receptorių.
Su visa savo įvairove GPCR yra monomeriniai vientisi membraniniai baltymai, kurių polipeptidinė grandinė septynis kartus kerta ląstelės membraną. Visais atvejais receptorių vieta, atsakinga už sąveiką su pirminiu signalu, yra lokalizuota išorinėje membranos pusėje, o sritis, kuri liečiasi su G baltymu, yra jos citoplazminėje pusėje.
Pasroviui esantį GPCR signalo perdavimo kaskados komponentą vaizduoja G baltymas. Aptikta apie 20 skirtingų G baltymų, tarp kurių visų pirma paminėtini G s ir C, kurie atitinkamai stimuliuoja ir slopina adenilato ciklazę; G q , aktyvinanti fosfolipazę C; Jutimo ląstelių G baltymai: fotoreceptorius - G t (transducinas), uoslės - G o jei ir skonio - G g.
C-baltymai yra heterotrimerai, susidedantys iš trijų tipų subvienetų: ct, (S ir y), tačiau natūraliomis sąlygomis paskutiniai du subvienetai veikia kaip vienas Ru kompleksas. Svarbiausia C-baltymų savybė yra guani surišimo centras ant jų a-subvieneto – nukleotidai: GDP ir GTP (139, 145 pav.).Jei GTP yra prijungtas prie C baltymo, tai tai atitinka jo aktyvuotą būseną.Jei GDP yra nukleotidų surišimo centre, tada ši forma atitinka neaktyvią baltymo būseną (79 pav.).
Pagrindinis įvykis perduodant signalą iš receptoriaus, kurį veikia pirminis signalas, į G baltymą yra tai, kad aktyvuotas receptorius katalizuoja BVP, susieto su G baltymu, mainus į aplinkoje esančią GTP. Šį BVP/GTP mainą G baltyme lydi trimerinės G baltymo molekulės disociacija į du funkcinius subvienetus: α subvienetą, kuriame yra GTP, ir Py kompleksą (139, 145 paveikslai).
Tada vienas iš šių funkcinių subvienetų, kuris priklauso nuo signalizacijos sistemos tipo, sąveikauja su efektoriniu baltymu, vaizduojamu fermento arba jonų kanalo. Dėl to atitinkamai kinta jų katalizinis aktyvumas arba joninis laidumas, o tai savo ruožtu lemia antrinio pasiuntinio (arba jono) citoplazmos koncentracijos pokyčius ir galiausiai inicijuoja vienokį ar kitokį ląstelių atsaką.
Efektoriniai baltymai signalizacijos sistemose, pavyzdžiui, GPCR-G-baltymo efektoriaus baltymas, gali būti adenilato ciklazė, kuri katalizuoja cAMP sintezę iš ATP; fosfolipazė C, kuri hidrolizuoja fosfatidilinozitolį, sudarydama DAT ir 1P3; fosfodiesterazė, kuri skaido cGMP į GMP; kai kurių tipų kalio ir kalcio kanalai.
Svarbu, kad signalo perdavimo metu G-baltymo efektoriaus baltymo receptorių kaskadoje pradinis išorinis signalas gali būti sustiprintas (pastiprintas) daug kartų. Taip nutinka dėl to, kad viena receptorių molekulė, būdama aktyvuotoje būsenoje (R*), sugeba paversti kelias G baltymo molekules į aktyvuotą formą (G*).
Pavyzdžiui, regėjimo kaskadoje rodopsino^C^ecGMP-fosfodiesterazė kiekvienai R* molekulei gali pagaminti kelis šimtus ar net tūkstančius Gt* molekulių, o tai reiškia, kad pirmajame 7?*-»G* kaskados etape Išorinio signalo stiprinimo koeficientas yra 10 2 -10 3 . Nors kitame kaskados etape (C*^efektoriaus baltymas) kiekviena G* molekulė sąveikauja tik su viena efektorinio baltymo molekule, signalas čia taip pat sustiprinamas, nes kiekvienai G* ir atitinkamai aktyvuotai molekulei. efektorinio baltymo, daugelis atsiranda (arba išnyksta) citoplazmoje antrinių pasiuntinių molekulių. Taigi, regėjimo kaskadoje antrajame etape viena aktyvuotos cGMP fosfodiesterazės molekulė gali suskaidyti iki 3000 cGMP molekulių per sekundę, kuri yra antrinis pasiuntinys fotoreceptorių ląstelėse.
Bendras kaskados padidėjimas yra lygus padidėjimo visuose kaskados etapuose sandaugai. Signalo, einančio per kaskadą, stiprinimo koeficientas gali pasiekti labai aukštas reikšmes: regos ląstelėse tai yra 10 5 -10 6 dydžio reikšmė.
Išorinio dirgiklio nutraukimą lydi visų signalizacijos sistemos komponentų išjungimas. Receptoriaus lygiu tai pasiekiama, pirma, dėl pirminio pasiuntinio atsiribojimo nuo komplekso su GPCR, ir, antra, fosforilinant receptorius, veikiant specialioms proteinkinazėms ir vėliau prisijungus prie modifikuoto specialaus receptoriaus. baltymų (pavyzdžiui, P-arrestino).
G-baltymai turi GTPazės aktyvumą, tai yra gebėjimą hidrolizuoti su jais susijusį GTP į BVP, o tai užtikrina jų savarankišką pašalinimą, tai yra G-GTP-e G-BVP perėjimą. Kadangi efektorinio baltymo aktyvacijos būsena (įjungta-išjungta) tiesiogiai priklauso nuo G-baltymo būsenos, šis perėjimas reiškia ir efektorinio baltymo išjungimą, taigi ir jo sintezės (hidrolizės) nutraukimą. antrinis pasiuntinys arba jonų kanalo uždarymas.
Ir galiausiai, kad ląstelės perėjimas į pradinę (prieš išorinį dirgiklį) būseną būtų baigtas, specialūs mechanizmai atkuria pradinį antrinio pasiuntinio arba katijono lygį jos citoplazmoje. Pavyzdžiui, cAMP, kurio citoplazminė koncentracija didėja perduodant signalą P-adrenerginių receptorių kaskadoje C5-baltymų-adenilato ciklazėje, cAMP fosfodiesterazės hidrolizuojasi į neciklinį (linijinį) AMP, kuris neturi savybių. antrinio pasiuntinio.
TRANSMEMBRANINIS SIGNALO PERDAVIMAS. Svarbi membranų savybė yra gebėjimas suvokti ir perduoti signalus iš išorinės aplinkos į ląstelę. Signalinių molekulių „atpažinimas“ atliekamas naudojant receptorių baltymus, įmontuotus į tikslinių ląstelių membraną arba esančius ląstelėje.
Jei signalą suvokia membraniniai receptoriai, informacijos perdavimo schema gali būti pavaizduota taip:
receptoriaus sąveika su signaline molekule (pirminiu pasiuntiniu);
membraninio fermento, atsakingo už antrojo pasiuntinio susidarymą, aktyvavimas;
antrinio pasiuntinio cAMP, cGMP, IP3, DAT arba Ca 2+ susidarymas;
specifinių baltymų, daugiausia baltymų kinazių, aktyvinimas, kuris, savo ruožtu, fosforilinant fermentus, įtakoja intracelulinių procesų aktyvumą.
Yra keletas transmembraninės informacijos perdavimo mechanizmų: naudojant adenilato ciklazės sistemą, inozitolio fosfato sistemą, katalizinius receptorius, citoplazminius arba branduolinius receptorius.
G-baltymų struktūrinė ir funkcinė organizacija
G-baltymai (GTP surišantys baltymai) yra universalūs tarpininkai, perduodantys signalus iš receptorių į ląstelės membranos fermentus, kurie katalizuoja antrųjų hormoninio signalo pasiuntinių susidarymą. G baltymai yra oligomerai, susidedantys iš α, β ir γ subvienetų.
Kiekvienas G baltymo α-subvienetas turi specifinius centrus:
GTP arba BVP susiejimas;
sąveika su receptoriumi;
prisijungimas prie βγ subvienetų;
fosforilinimas proteinkinaze C;
sąveika su fermentu adenilato ciklaze arba fosfolipaze C.
G baltymų struktūroje trūksta α-spiralinių, membraną apimančių domenų. G baltymai priklauso „įtvirtintų“ baltymų grupei.
G-baltymų aktyvumo reguliavimas
Yra neaktyvi G baltymo forma – αβγ-BVP kompleksas ir aktyvuota forma αβγ-GTP. G baltymo aktyvacija įvyksta sąveikaujant su aktyvatoriaus-receptoriaus kompleksu; G baltymo konformacijos pasikeitimas sumažina α-subvieneto afinitetą BVP molekulei ir padidina jį GTP. BVP pakeitimas GTP aktyviojoje G baltymo vietoje sutrikdo α-GTP ir βγ subvienetų komplementarumą. Receptorius, susijęs su signaline molekule, gali suaktyvinti daugybę G baltymo molekulių, taip užtikrindamas ekstraląstelinio signalo amplifikaciją šiame etape.
Aktyvuotas G baltymo α-subvienetas (α-GTP) sąveikauja su specifiniu ląstelės membranos baltymu ir keičia jo aktyvumą. Tokie baltymai gali būti fermentai adenilato ciklazė, fosfolipazė C, cGMP fosfodiesterazė, Na+ kanalai, K+ kanalai.
Ryžiai. 5-35. G-baltymų funkcionavimo ciklas. R s – receptorius; G – hormonas; AC – adenilato ciklazė.
Kitas G-baltymo funkcionavimo ciklo etapas yra GTP, prijungto prie α-subvieneto, defosforilinimas, o fermentas, katalizuojantis šią reakciją, yra pats α-subvienetas.
Dėl defosforilinimo susidaro α-BVP kompleksas, kuris nėra papildomas specifiniam membranos baltymui (pavyzdžiui, adenilato ciklazei), tačiau turi didelį afinitetą py protomerams. G baltymas grįžta į neaktyvią formą – αβγ-BVP. Vėliau suaktyvinus receptorių ir pakeitus GDP molekulę GTP, ciklas kartojasi dar kartą. Taigi, G baltymų α-subvienetai perneša stimuliuojantį arba slopinamąjį signalą iš receptoriaus, kurį aktyvuoja pirminis pasiuntinys (pvz., hormonas), į fermentą, katalizuojantį antrinio pasiuntinio susidarymą.
Kai kurios baltymų kinazių formos gali fosforilinti G baltymų α-subvienetus. Fosforilintas α-subvienetas nėra papildytas specifiniu membranos baltymu, tokiu kaip adenilato ciklazė arba fosfolipazė C, todėl negali dalyvauti signalo perdavime.
Adenilato ciklazė
Fermentas adenilato ciklazė, kuris katalizuoja ATP pavertimą cAMP, yra pagrindinis fermentas adenilato ciklazės signalo perdavimo sistemoje. Fermentas priklauso integralinių ląstelės membranos baltymų grupei, turi 12 transmembraninių domenų. Ekstraląsteliniai adenilato ciklazės fragmentai yra glikozilinti. Adenilato ciklazės citoplazminiai domenai turi du katalizinius centrus, atsakingus už cAMP, antrojo pasiuntinio, dalyvaujančio reguliuojant baltymo kinazės A fermento aktyvumą, susidarymą.
Adenilato ciklazės aktyvumui įtakos turi tiek tarpląsteliniai, tiek tarpląsteliniai reguliatoriai. Ekstraląsteliniai reguliatoriai (hormonai, eikozanoidai, biogeniniai aminai) reguliavimą vykdo per specifinius receptorius, kurie, naudodami G baltymų α-subvienetus, perduoda signalus į adenilato ciklazę. α s-subvienetas (stimuliuojantis), sąveikaudamas su adenilato ciklaze, aktyvina fermentą, α-subvienetas (slopinantis) slopina fermentą. Iš 8 tirtų adenilato ciklazės izoformų 4 yra priklausomos nuo Ca 2+ (aktyvuojamos Ca 2+). Adenilato ciklazės reguliavimas viduląsteliniu kalciu leidžia ląstelei integruoti dviejų pagrindinių antrųjų pasiuntinių cAMP ir Ca 2+ veiklą.
Adenilato ciklazės sistema
Dalyvaujant adenilato ciklazės sistemai, realizuojamas šimtų skirtingo pobūdžio signalinių molekulių – hormonų, neurotransmiterių, eikozanoidų – poveikis.
Transmembraninės signalizacijos sistemos funkcionavimą užtikrina baltymai: signalinės molekulės Rs-receptorius, aktyvinantis adenilatciklazę, ir signalinės molekulės R i -receptorius, slopinantis adenilatciklazę; G s -stimuliuojantys ir G j -adenilato ciklazę slopinantys baltymai; fermentai adenilato ciklazė (AC) ir proteinkinazė A (PKA).
Įvykių, dėl kurių suaktyvėja adenilato ciklazė, seka:
adenilato ciklazės sistemos aktyvatoriaus, pavyzdžiui, hormono (G) prisijungimas prie receptoriaus (R s), sukelia receptoriaus konformacijos pasikeitimą ir jo afiniteto Gs baltymui padidėjimą. Dėl to susidaro kompleksas [G][R][O-BVP];
[G][R] pridėjimas prie G-BVP sumažina Gs baltymo α-subvieneto afinitetą BVP ir padidina giminingumą GTP. BVP pakeičiamas GTP;
tai sukelia komplekso disociaciją. Atskirtas α subvienetas, susietas su GTP molekule, turi afinitetą adenilato ciklazei:
[G][R] → [G][R] + α-GTP + βγ;
α-subvieneto sąveika su adenilato ciklaze lemia konformacijos pokyčius
fermentas ir jo aktyvinimas, padidėja cAMP susidarymo iš ATP greitis;
Konformaciniai [α-GTP][AC] komplekso pokyčiai skatina α-subvieneto GTP-fosfatazės aktyvumo padidėjimą. Vyksta GTP defosforilinimo reakcija, ir vienas iš reakcijos produktų – neorganinis fosfatas (Pi) – atskiriamas nuo α-subvieneto ir išlaikomas [α-BVP] kompleksas; hidrolizės greitis lemia signalo perdavimo laiką;
BVP molekulės susidarymas aktyviame α-subvieneto centre sumažina jos afinitetą adenilato ciklazei, bet padidina afinitetą βγ-subvienetams. G s baltymas grįžta į neaktyvią formą;
jei receptorius yra susijęs su aktyvatoriumi, pavyzdžiui, hormonu, G baltymo funkcionavimo ciklas kartojamas.

Adenilato ciklazės sistema
Svarbiausias tarpląstelinis signalizacijos kaskadų komponentas yra G baltymai. Šiuo metu žinoma apie 20 skirtingų G baltymų. Pavyzdžiui, Gs ir Gi atitinkamai stimuliuoja ir slopina adenilato ciklazę; Gq aktyvina fosfolipazę C. Tarp jutimo ląstelių G baltymų galime pastebėti: fotoreceptorių – Gt (transducin), uoslės – Golf ir skonio – Gg.
Pagal savo struktūrą G baltymai yra heterotrimeriai, susidedantys iš trijų tipų subvienetų: a (alfa), b (beta) ir g (gama), tačiau natūraliomis sąlygomis beta ir gama subvienetai veikia kaip vienas kompleksas. Bendras G baltymų struktūrinis bruožas yra septynių transmembraninių alfa spiralių buvimas. Svarbiausia G baltymų savybė yra tai, kad jų α-subvienete yra guanilo nukleotidų surišimo centras: GDP (guanisidino difosfatas) ir GTP (guanisidino trifosfatas). Jei GTP yra prijungtas prie G baltymo, tai atitinka jo aktyvuotą būseną (G-GTP) arba, kitaip, G baltymas yra aktyvuotoje padėtyje. Jei BVP yra nukleotidų surišimo vietoje, tada ši forma (G-BVP) atitinka „išjungimo“ būseną. Pagrindinis signalo perdavimo iš receptoriaus (kurį paveikė pirminis signalas) į G baltymą taškas yra aktyvuoto receptoriaus katalizė, kai BVP, susietas su G baltymu, keičiasi į aplinkoje esančią GTP (BVP / GTP). mainai G baltymu).
Transmembraniniai receptoriai atlieka pagrindines ląstelės gyvybines funkcijas: signalizaciją, transportavimą, apsaugą. Įvairių biologiškai aktyvių junginių, įskaitant antivirusinius ir antibakterinius, veikimo mechanizmo tyrimas parodė, kad specifiškiausi tiek vaistinių, tiek toksinių junginių (nuodų) taikiniai yra žmogaus ląstelių receptoriai ir patogeniniai mikroorganizmai. Didelė dalis transmembraninių receptorių yra G baltymų sujungti receptoriai (GPCR), Maždaug pusė visų šiuo metu žinomų vaistų veikia būtent GPCR. Iš visų ląstelių paviršiaus receptorių tipų GPCR yra universaliausi. Šie receptoriai suriša daugybę molekulių – nuo mažų neurotransmiterių iki didelių baltymų. GPCR dalyvauja beveik visuose gyvybiškai svarbiuose procesuose.
GPCR perduodamų signalų įvairovę užtikrina funkcinis skirtingų GPCR sujungimas tarpusavyje. Taigi akivaizdu, kad universaliausias toksinių ir vaistinių junginių poveikio ląstelei mechanizmas realizuojamas per poveikį ląstelės receptorių aparatui, keičiant jų konformaciją arba pagrindines ligando-receptoriaus ryšio charakteristikas, jų specifiškumą ir grįžtamumas.

2018 metais gimimo pasninkas prasidės lapkričio 28 d. Šiuo laikotarpiu stačiatikiai ruošiasi švęsti Kalėdas...

Daugumos moterų svajonė yra sukurti šeimą. Jie nori turėti mylintį vyrą ir krūvą vaikų. Bet tai ne visada yra santykiai...

Šiame straipsnyje pateikiama: galingiausia skyrybų malda – informacija, paimta iš viso pasaulio, elektroninio tinklo ir...

Informacinė svetainė apie ikonas, maldas, stačiatikių tradicijas. Malda už skandalus ir kivirčus šeimoje, su vyru, su vaikais...

Kokie būtų Naujieji metai be šampano, mandarinų, Olivier, aspicų ir visų mėgstamo „Silkės po kailiu“. Su paskutiniu...

Paruoškime sausainiams reikalingus ingredientus. Pirmas dalykas, kurį reikia padaryti, yra užvirti vandenį. Mums reikia...

Ar galima registruoti darbuotoją į finansų direktoriaus – vyriausiojo buhalterio pareigas? Vyriausioji buhalterė tvirtina...

Smulkaus verslo vadovas gali lengvai valdyti biudžetą savarankiškai. PATIKRINTA! Jei pavyks...

Naujų projektų kūrimas apima preliminarų ekonominį jų pagrįstumo pagrindimą, vėlesnius...
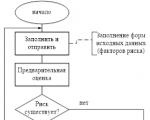
Ataskaitas rengia RM, dėl jų susitaria (patvirtina) Rizikos komitetas prie valdybos ir perduoda...

Didelės, labai didelės pievos pakraštyje, ant ilgo smaragdinio žolės stiebo gyveno mažytė boružėlė. Dievo mažutė...

Šiais laikais gana įprasta, kad žmonės atsigręžia į žvaigždes. Horoskopo pagalba žmogus gali sužinoti...

Verslo ar draugiško. Jei persekiojote nepažįstamą žmogų, tai reiškia jūsų pasitikėjimo moteriškąja lytimi lygį...

pagal Freudo svajonių knygą Jei svajojote, kaip žvejojote, tai reiškia, kad realiame gyvenime vargu ar galite išsijungti...

Daugumos moterų svajonė yra sukurti šeimą. Jie nori turėti mylintį vyrą ir krūvą vaikų. Bet ne visada...

Šiame straipsnyje yra: pati galingiausia malda už skyrybas – informacija paimta iš viso pasaulio, elektroninė...