Spanish Fly for two – hogyan befolyásolják a libidót nőkben és férfiakban
Tartalom: Biológiailag aktív adalékanyag, amely egy légyes bogárból (vagy légy...
A sejtváz - a citoszkeleton - tanulmányozásának szentelt rész
A mikrotubulus felezési ideje ~5 perc, a mitózis első felében ~15s
A mikrotubulus átmérője 25 nm.
A mikrotubulus szerkezeti egysége egy fehérje heterodimer tubulin, α- és β-alegységekből áll (53 és 55 kDa), külön nem érkezik, hasonló, de nem azonos. Mindegyik alegységnek van egy nukleotidkötő helye. Az α-tubulin megköti a GTP molekulát, amely nem hidrolizálódik, a β-tubulin képes megkötni a GDP-t vagy a GTP-t (1. ábra). Az egyik heterodimer β-tubulinja megköti a GTP-t és összekapcsolódik egy másik heterodimer α-tubulinjával, míg a GTP GDP-vé hidrolizálódik. Az α-tubulin egy GTP-aktiváló fehérje, és katalizálja a β-tubulin GTP hidrolízisét (2. ábra). Így a heterodimerek lineáris láncokat alkotnak - protofilamentumokat, 13 protofilamentum helikális ciklusos komplexet alkot, az ilyen gyűrűk csővé polimerizálódnak (3. ábra). A tubulin foszforilációja fokozza a polimerizációt.
1. ábra Tubulin heterodimer. α-tubulin (syn.) GTP-kötőhellyel (kék). β-tubulin (zöld) GTP- és GDP-kötő helyekkel (piros)
A mikrotubulusok dinamikus poláris str-ryek. A (+)-terminális dinamikusan instabil (β-tubulin), a (-)-terminális pedig a mikrotubulus-szervező központhoz való kötődés révén stabilizálódik (lásd a Centrosome áttekintését).
A menetmarás a mikrotubulusok mozgása a mikrotubulusok egyik végének egyidejű növekedése és a másik végének disszociációja következtében.
A nukleotid-kötő doménben lévő tubulin DNS-nek erősen konzervált GGGTG(T/S)G szekvenciája van.
Az FtsZ bakteriális fehérje, a tubulin homológja, a bakteriális citoszkeleton összetevője, és polimerizálva mikrotubulusokat képez.
 mikrotubulusok
mikrotubulusok
2. ábra A mikrotubulusok képesek szingulettet, dublettet és triplettet alkotni.
A dublett vagy triplett mikrotubulusa 13 protofilamentumból áll.
A B és C tubulusok kevesebb protofilamentumból állnak, jellemzően 10-ből.
A mikrotubulusokhoz kötődő fehérjék.
A mikrotubulusokhoz kétféle fehérje kapcsolódik: szerkezeti
fehérjék (MAP-microtubuls-asssociated proteins) és transzlokátor fehérjék.
A MAP kötődést foszforiláció szabályozza, aminek eredményeként
amelyeket egyes MAP-ok leválasztanak a mikrotubulusokról.
+TIPPEK- a (+) véggel kölcsönhatásba lépő fehérjék
mikrotubulusok, amelyek közül sok motorfehérje,
mások interakciót biztosítanak a mikrofilamentumokkal
sejtkéreg mikrotubulusok plazmához erősítésével
membrán. Néhány +TIPS szabályozza a mikrotubulusok dinamikáját
és a (+) vég stabilitása, például, XMAP215
A fehérjék családja stabilizálja a (+)-terminálist, megakadályozva a degradációt
és lehetővé teszi a mikrotubulusok növekedését.
KAPOCS- kötőfehérjék
tubulin dimerek a (+)-végig és gátolják katasztrófák.
Kölcsönhatásba lépnek a kinetochore-val – egy komplex, amely összeköt
a mikrotubulus (+)-vége a kromoszómával.
Katasztrófák - +TIP fehérjék, amelyek a mikrotubulusok (+) végéhez kötődnek
és a tubulin dimerek disszociációját biztosítja. Képesek
aktiválja a GTP hidrolízist vagy a protofilamentum konformáció változását
(MCAK- a kinezin a kinetokorban található
és biztosítja a (+)-vég disszociációját a mitózis anafázisában).
Stasmin- destabilizáló fehérje
rákos sejtekben. Tubulin heterodimerrel kötődik
akadályozzák polimerizációjukat. A sztazminokat a foszforiláció gátolja.
Katanin - elválasztja a mikrotubulusokat, hogy új instabilt képezzen
(+)-vég.
Egyes MAP-ok mikrotubulusokat kötnek össze
egymással, membránnal vagy közbenső szálakkal.
Az I. típusú MAP az idegsejtek axonjaiban és dendritjeiben található
és néhány másikban a KKEX többszörös ismétlődése (Lys-Lys-Glu-X)
amelyek megkötik a tubulin (-) töltött szakaszait.
A II-es típusú MAP megtalálható az idegek axonjaiban és dendriteiben is
sejtek és néhány más. 3-4 ismétlődésük van 18 maradékból
szekvencia, amely a tubulint köti.
A mikrotubulusok (+) végével kölcsönhatásba lépő fehérjék
APC, Kar9 ( sc)*
APC (adenomatous polyposis coli) - tumorszuppresszor,
amely a fehérje komplex szabályozásának alapja
b-kateninek foszforilációja.
EB1, Bim1(sc) , Mal3(sp)
EB1 (végkötő fehérje 1) - fehérje kölcsönhatásba lép
APC.
meztelen(an)
Nud (nukleáris eloszlás) - a dyneineket szabályozó fehérje.
Lis1/NUDF(An) Pac1(sc)
Lis (lissencephaly) - az emberi agy fejlődésének megsértése
(sima agy). A fehérje kölcsönhatásba lép a dyneinnel a szabályozás érdekében
a funkciója.
MEZTELEN(An) R011(Neurospóra
crassa) /Ndl1(sc) ; Nde1, Ndel1
(emlősök).
Ezek a fehérjék kölcsönhatásba lépnek a Lis1-gyel és a deneinekkel, és biztosítják
működésüket.
Kar3(sc)
A Kar3 egy kinezin, amelynek C-terminális motordoménje van, és ehhez tartozik
a Kinesin-14 családnak.
Kip2(sc), Tea 2
(Sp), KipA(An)
A Kinesin-7 családba tartozó gombás kinezinek, beleértve
CENP-E - emlős centromer fehérje, Kip2 , Tea2 és
KipA
Klp10A(Dm), Klp59C, MCAK
A Kinesin-13 család tagjai. Klp10A – feltételezett homológ
Kif2A emlősök. Klp59C (Dm) – feltételezett homológ
MCAK emlősök. KLP10A és más tagok I. rokon
a kinezinek alcsaládjai, amelyek kölcsönhatásba lépnek a fedetlennel
A hasadási orsó mikrotubulusainak (-)-vége a mitózis során.
Biztosítják a tubulin pólus dimerek disszociációját
sejteket, hozzájárulva tedmilling(mozgalom
mikrotubulusok a pólusokhoz és a mikrotubulusok lerövidülése közben
mitózis anafázisa).
Dynactin
Fehérje komplex, beleértve a p150 ragasztott fehérjét. Dinaktin köt
dyneint és szabályozza annak tulajdonságait, valamint hólyagokat is rögzít
a dyneinhez. A p150glued egy NUDMA homológ. nidulans.
CLIP-170, Bik1 (sc), Tipp
(Sp)
A CLIP-170 biztosítja a mikrotubulusok stabilizálását és növekedését,
és szabályozza a dynein lokalizációját is.
CLIP-170 - biztosítja a dynein-dynactin komplex leszállását,
részt vesz a hólyagok szállításában, a mikrotubulus végén.
A LIP-170 a citoplazmában inaktív konformációban található
amelyben a mikrotubulus-kötő N-terminális kapcsolódik
ugyanazon molekula C-terminálisával. Amikor az N-terminálist tubulinhoz köti
vagy a mikrotubulus (+)-vége, a C-terminális felszabadul és megköti
dynein-dynactin komplexszel p150-en keresztül Ragasztott molekula, mikrotubulus
stabilizálja. Dynenin-dynactin felszabadul és elkezdődik
mozgás a mikrotubulus mentén (3. ábra) 
Egyes toxinok és gyógyszerek, amelyek némelyike zavarja a mitózist, zavarja a tubulin polimerizációját és depolimerizációját:
A Taxol egy rákellenes gyógyszer, amely stabilizálja a mikrotubulusokat.
A kolhicin a polimerizáció blokkolásával köti meg a tubulint. A mikrotubulusok depolimerizálódnak magas kolhicinkoncentráció esetén.
vinblasztin - fokozza a depolimerizációt azáltal, hogy vinblasztin-tubulin parakristályokat képez.
nokodazol - biztosítja a mikrotubulusok depolimerizációját.
Az asszociációt elnyomja a vinblasztin, vinkrisztin, kolhicin, és fokozza a taxol.
A gamma-szóma a mikrotubulusokat szervező központ a sejtmag külső felületén.
G-aktin monomer (globuláris aktin) - aszimmetrikus
(42 kDa) két doménből áll, mint az ionos
az aggregátumokat egy tekercselt polimer F-aktinná (fibrilláris
aktin).
A G-aktin kötőhelyekkel rendelkezik a kétértékű kationokhoz
és nukleotidok Mg 2+ által elfoglalt fiziológiás körülmények között
és ATP.
A G-aktin polimerizációja F-aktinná
Az F-aktin polaritással (+) és (-) rendelkezik
különféle tulajdonságok.
A G-aktin molekula szorosan kötött ATP-t hordoz, amely, amikor
az F-aktinná való átmenet lassan ADP-vé hidrolizál - mutat
Az ATPáz tulajdonságai A polimerizációt hidrolízis kíséri
ATP, ami nem szükséges, mert jelenlétében megy végbe a polimerizáció
az ATP nem hidrolizálható analógjai
A polimerizáció több folyamatból áll: magképződés,
megnyúlás, disszociáció,
töredezettség, dokkolás.
Ezek a folyamatok egyszerre futnak.
Nukleáció- három G-aktin kapcsolata
polimerizáció beindítása.
Megnyúlás- az aktinlánc meghosszabbítása a
a G-aktin kapcsolódása az F-aktin (+)-terminálisához.
Disszociáció- lánc rövidítés. Depolimerizáció
Az aktin mindkét végén azonos sebességű
Töredezettség- hőmozgás hatására
F-aktin fragmentálhat.
Dokkolás- külön töredékek kapcsolhatók össze
egymással a végétől a végéig.
G>F koncentrációnál a polimerizáció egyidejűleg megy végbe
(+) és (-) vége.
Ha G
a (+)-vég egyidejű kiterjesztése és a disszociáció miatt
(-)-vége. G ~ F-nél - dinamikus egyensúly - következik be
a (+) vég polimerizálása és a (–) vég depolimerizálása költséggel
energia ATP A G-aktin kötődik az ATP-hez és polimerizál, hidrolizál
ATP. a G-aktin kritikus végein (+), a vége meghosszabbodik,
és (-) - rövidítve
aktin mikrofilamentumok
F-aktin - fibrilláris, hélix fordulat hossza 37
nm, d=6-8 nm.
Aktinkötő fehérjék
A citoplazmában több mint 50 fehérje kötődik aktinhoz
különböző funkciók: szabályozza a G-aktin medence (profilin) mennyiségét,
befolyásolja a polimerizáció sebességét (villine), stabilizálja
a szálak végeit (fragin, a-actinin), varrjuk le a szálakat
más vagy más komponensekkel (villin, α-aktin, spektrin,
MARCKS, fimbrin), elpusztítja az F-aktin (gelsolin) kettős hélixét.
Ezen fehérjék aktivitását a Ca 2+ és a protein kinázok szabályozzák.
A fehérjéknek öt helye van: a monomerrel
aktin, (+)-véggel (tollas), (-)-véggel (hegyes),
oldalfelülettel. Az aktinkötő fehérjék lehetnek
Ca 2+ -ra érzékeny vagy érzéketlen
1. Az aktin monomerhez kötődő fehérjék - gátolják a magképződést
(profilin, fragmentin - Ca 2+ -ra érzékeny).
A profilin monomerrel képes F-aktint, míg a fragmentint felépíteni
nem, blokkolja mind a magképződést, mind a megnyúlást. Nem érzékeny
Ca 2+ DNáz I-re és vitaminkötő fehérjére
D - funkció a sejten kívül.
2. A záró (+)-vég zárolható sapkával
fehérjék - blokkolják a megnyúlást és a dokkolást, hozzájárulnak
gócképződés - lerövidült filamentumok megjelenése (gelsolin,
villin, fragmin)
3. (-)-vég - magképződés megindítása, dokkolás elnyomása
és megnyúlás - a töredékek számának növekedése és hosszának csökkenése.
Acumentin makrofágokban, brevin - tejsavófehérje okoz
az F-aktin oldat viszkozitásának gyors csökkenése. Mindkét fehérje
Ca 2+ -ra érzékeny
4. Nem keresztkötés – az oldalsó kötés egyaránt stabilizálódhat
és destabilizálja az F-aktin tropomiozint (Ca-független)
stabilizálja, szeverin, villin (Ca-függő) - kötődés
F-aktinnal vágja le.
5. F-aktin térhálósodás gélképződéssel. Ilyen
fehérjék nukleációt indukálnak. Az ilyen fehérjék dimerek vagy rendelkeznek
két aktinkötő domén. vérlemezke α-aktin,
villin, fimbrin, aktinogelin makrofágokból (Ca-független).
capping fehérjék- zárja le az aktin végeit
filamentek, megakadályozzák a polimerizációt-depolimerizációt,
elősegítik az izzószálnak a membránhoz való tapadását.
falloidin- a sápadt gombagomba mérge, megköti
a (-) véggel és gátolja a depolarizációt.
citokalazin– penészméreg csatlakozik
a (+) vég felé, blokkolva a polimerizációt.
záró-fragmentáló fehérjék- töredék
F-aktin gélből szolba átalakulást okoz (Gelsolin 90 kD aktiválva
A Ca2+ 10-6M megszakítja az F-aktint és megköti a végét).
F-aktint kötő fehérjék
| fehérje | M, kD | rizs. | lokalizáció és az F-aktinra gyakorolt hatás |
| fascin | 55 | phyllopodia, lamelopodia, stresszrostok, mikrobolyhok, akroszóma |
|
| tropomiozin | 2x35 | stabilizálja az F-aktint, megakadályozza a fragmentációt | |
| miozin | 2x260 | cérnacsúszás | |
| minimiozin | 150 | buborékmozgás | |
| profilin | 15 | G-aktin tárolása | |
| átvizsgálja | 102 | akroszóma | |
| viline | 92 | mikrobolyhok | |
| dematin | 48 | eritrociták kérgi hálózata | |
| fimbrin | 68 | tapadás. érintkezők, mikrovillák kötegben | |
| aktinin | 2x102 | adhéziós érintkezők, nyalábokba kötött mikrobolyhok | |
| spektrin | 2x265+2x260 | vörösvértestek tőrhálózata, a PM-hez való kapcsolódás | |
| disztrofin | 427 | cor.net egérszálak | |
| ABP120 | 92 | pszeudopodia | |
| filamin | 2x280 | pszeudopodia, a stresszszálak hálózatba varrnak össze |
Aktin által alkotott szerkezetek
Sejtes kéreg- aktinszálak hálózata
a plazmamembrán alatt.
Phyllopodium
Stresszrostok – akkor keletkeznek, amikor egy sejt rendelkezik
az aljzathoz való rögzítési képesség
KÖZBETŰ FILAMENTEK
sejtfilamentumok közötti fehérjék száma M, kD típus
savas keratin epit >15 40-57 I
alap keratin epit >15 53-67 II
desmin egér 1 53 III
savas fibrilláris fehérje glia, asztrociták 1 50
vimentin mesenkh, nyaki epit 1 57
perifériás idegrendszer 1 57
neurofilamentum fehérjék: axonok és dendritek IV
NF-L 162
NF-M 1 102
NF-H 1110
internexin CNS 1 66
nestin epit idegszövet 1 240
Az összes sejt A sejtmagja 1 70 V
Lamin B 1 67
laminált C 1 67
szeptamer monomer, párhuzamos dimer, antiparallel tetramer, protofilamentum protofibrill?PF
közbenső szálak
d=10nm, (citokeratinek, dezmin, vimentin, savas fibrilláris gliaprotein (GFAP), neurofilamentum) egy bázikus rúdból áll - egy szuperspirálos -hélix, az ilyen dimerek antiparallel asszociálnak, tetramert képeznek, tetramerek aggregációja "fejtől-fejig" " protofilamentumot ad, 8 protofilamentes képet. köztes szál | polimerizáció képhez vezet. stabil apoláris polimer molekulák
IF-asszociált fehérjék
protein M, kD lokalizáció
BPAG1 230 hemidesmoszóma
placoglobin 3 dezmoszómák
dezmoplacin I 250 desm
desmoplakin II 215 desm
plectin 300 cortec. zóna
ankyrin 140 cortec. zóna
filaggrin 30 citoszol
B-lamin receptor 58 mag
A mutáns egerekből hiányzik a vimentin, míg az egerek teljesen normálisan élnek.
A növényi sejtekben a citoszkeletont mikrotubulusok és mikrofilamentumok képviselik, köztes filamentumok nincsenek, laminák viszont vannak.
Szempilla - citoplazma kinövése h=300nm, pm borítja
axoneme – d=200nm, 9 dupla mikrotubulus, 100, 2 centrális mikrotubulus, A-mikrotubulus - 13 alegység, B-mikrotubulus - 11 alegység,
alaptest - a citoplazmába merülve d = 200 nm, 9 mikrotubulus hármas, fogantyúi, hüvelye és küllői vannak a proximális részén.
A csillók miatti sejtmozgás sebessége elérheti a ~5 mm/s-ot. A légcső sejtjében a csillók száma ~300, a csillófélék sejtjében ~14 ezer.
kinetocylium - mozgásképes (hám, spermium), elsődleges csillók - nem mozognak. 
A Wikipédiából, a szabad enciklopédiából
eukarióta citoszkeleton. Az aktin mikrofilamentumok vörösre, a mikrotubulusok zöldre, a sejtmagok kékre festődnek.
Citoszkeleton- ez egy sejtváz vagy váz, amely egy élő sejt citoplazmájában található. Minden eukarióta sejtben jelen van, és az összes eukarióta citoszkeletális fehérje homológját megtalálták a prokarióta sejtekben. A citoszkeleton egy dinamikus, változó struktúra, melynek feladata a sejt alakjának megtartása és hozzáigazítása a külső hatásokhoz, exo- és endocitózishoz, biztosítva a sejt egészének mozgását, az aktív intracelluláris transzportot és sejtosztódást.

Keratin közbenső filamentumok egy sejtben.
A citoszkeletont fehérjék alkotják, több fő rendszer létezik, amelyeket vagy az elektronmikroszkópos vizsgálatok során látható fő szerkezeti elemek (mikrofilamentumok, köztes filamentumok, mikrotubulusok), vagy az összetételüket alkotó fő fehérjék (aktin-miozin) alapján neveznek el. rendszer, keratinok, tubulin – dynein rendszer).
A körülbelül 7 nm átmérőjű mikrofilamentumok két spirális aktinmonomer lánc. Főleg a sejt külső membránján koncentrálódnak, mivel a sejt alakjáért felelősek, és képesek a sejtfelszínen kiemelkedéseket (pszeudopodiákat és mikrobolyhokat) kialakítani. Részt vesznek az intercelluláris interakcióban (tapadó kontaktusok kialakításában), a jelátvitelben és a miozinnal együtt az izomösszehúzódásban is. A citoplazmatikus miozinok segítségével mikrofilamentumok mentén vezikuláris transzport valósítható meg.
Sokáig azt hitték, hogy csak az eukariótáknak van citoszkeletonja. Azonban Jones és munkatársai 2001-es tanulmánya óta. (PMID 11290328), amely leírja a bakteriális aktin homológok szerepét a sejtekben Bacillus subtilis, megkezdődött a bakteriális citoszkeleton elemeinek aktív tanulmányozásának időszaka. A mai napig mindhárom típusú eukarióta citoszkeletális elem – a tubulin, az aktin és a köztes filamentumok – bakteriális homológjait megtalálták. Azt is megállapították, hogy a bakteriális citoszkeleton fehérjék legalább egy csoportja, a MinD/ParA, nem rendelkezik eukarióta analógokkal.
A citoszkeleton leginkább vizsgált aktinszerű komponensei az MreB, a ParM és a MamK.
Az MreB fehérjék és homológjai a bakteriális citoszkeleton aktinszerű komponensei, amelyek fontos szerepet játszanak a sejtforma fenntartásában, a kromoszóma szegregációban és a membránszerkezetek szerveződésében. Egyes baktériumfajták, mint pl Escherichia coli, csak egy MreB fehérjét tartalmaznak, míg másokban 2 vagy több MreB-szerű fehérje található. Ez utóbbira példa a baktérium Bacillus subtilis, amelyben MreB fehérjék, Mbl ( Mújra B-l ike) és MreBH ( MreB h omolog).
A genomokban E. coliÉs B. subtilis az MreB szintéziséért felelős gén ugyanabban az operonban található, mint az MreC és MreD fehérjék génjei. Az ezen operon expresszióját elnyomó mutációk csökkent életképességű gömbsejtek kialakulásához vezetnek.
Az MreB fehérje alegységei szálakat alkotnak, amelyek egy pálcika alakú baktériumsejt köré tekerednek. A citoplazma membrán belső felületén helyezkednek el. Az MreB által alkotott filamentumok dinamikusak, folyamatosan polimerizálódnak és depolimerizálódnak. Közvetlenül a sejtosztódás előtt az MreB azon a területen koncentrálódik, ahol a szűkület kialakul. Úgy gondolják, hogy az MreB funkciója a murein, egy sejtfal polimer szintézisének koordinálása is.
Az MreB homológok szintéziséért felelős gének csak rúd alakú baktériumokban voltak megtalálhatók, coccusokban nem.
A ParM fehérje alacsony kópiaszámú plazmidokat tartalmazó sejtekben van jelen. Feladata a plazmidok hígítása a sejt pólusai mentén. Ugyanakkor a fehérje alegységei a pálcika alakú sejt nagy tengelye mentén kifeszített filamentumokat alkotnak.
Az izzószál szerkezetében kettős spirál. A ParM által alkotott filamentumok növekedése mindkét végén lehetséges, ellentétben az aktin filamentumokkal, amelyek csak a ±póluson nőnek.
A MamK egy aktinszerű fehérje Magnetospirillum magneticum felelős a magnetoszómák helyes elhelyezkedéséért. A magnetoszómák a vasrészecskéket körülvevő citoplazma membrán invaginációi. A MamK filamentum vezetőként működik, amely mentén a magnetoszómák egymás után helyezkednek el. A MamK fehérje hiányában a magnetoszómák véletlenszerűen oszlanak el a sejtfelszínen.
A Wikipédiából, a szabad enciklopédiából
Citoszkeleton- ez egy sejtváz vagy váz, amely egy élő sejt citoplazmájában található. Minden eukarióta sejtben jelen van, és az összes eukarióta citoszkeletális fehérje homológjait megtalálták a prokarióta sejtekben. A citoszkeleton egy dinamikus, változó struktúra, melynek feladata a sejt alakjának megtartása és hozzáigazítása a külső hatásokhoz, exo- és endocitózishoz, biztosítva a sejt egészének mozgását, az aktív intracelluláris transzportot és sejtosztódást.  A citoszkeletont fehérjék alkotják, több fő rendszert különböztetnek meg, amelyeket vagy az elektronmikroszkóppal látható fő szerkezeti elemek (mikrofilamentumok, köztes filamentumok, mikrotubulusok), vagy az összetételüket alkotó fő fehérjék (aktin-miozin rendszer, keratinok) alapján neveznek el. , tubulin - dynein rendszer).
A citoszkeletont fehérjék alkotják, több fő rendszert különböztetnek meg, amelyeket vagy az elektronmikroszkóppal látható fő szerkezeti elemek (mikrofilamentumok, köztes filamentumok, mikrotubulusok), vagy az összetételüket alkotó fő fehérjék (aktin-miozin rendszer, keratinok) alapján neveznek el. , tubulin - dynein rendszer).
A körülbelül 7 nm átmérőjű mikrofilamentumok két spirális aktinmonomer lánc. Főleg a sejt külső membránján koncentrálódnak, mivel a sejt alakjáért felelősek, és képesek a sejtfelszínen kiemelkedéseket (pszeudopodiákat és mikrobolyhokat) kialakítani. Részt vesznek az intercelluláris interakcióban (tapadó kontaktusok kialakításában), a jelátvitelben és a miozinnal együtt az izomösszehúzódásban is. A citoplazmatikus miozinok segítségével mikrofilamentumok mentén vezikuláris transzport valósítható meg.
Sokáig azt hitték, hogy csak az eukariótáknak van citoszkeletonja. Azonban Jones és munkatársai 2001-es tanulmánya óta. (), leírja a bakteriális aktin homológok szerepét a sejtekben Bacillus subtilis, megkezdődött a bakteriális citoszkeleton elemeinek aktív tanulmányozásának időszaka. A mai napig mindhárom típusú eukarióta citoszkeletális elem – a tubulin, az aktin és a köztes filamentumok – bakteriális homológjait megtalálták. Azt is megállapították, hogy a bakteriális citoszkeleton fehérjék legalább egy csoportja, a MinD/ParA, nem rendelkezik eukarióta analógokkal.
A citoszkeleton leginkább vizsgált aktinszerű komponensei az MreB, a ParM és a MamK.
Az MreB fehérjék és homológjai a bakteriális citoszkeleton aktinszerű komponensei, amelyek fontos szerepet játszanak a sejtforma fenntartásában, a kromoszóma szegregációban és a membránszerkezetek szerveződésében. Egyes baktériumfajták, mint pl Escherichia coli, csak egy MreB fehérjét tartalmaznak, míg másokban 2 vagy több MreB-szerű fehérje található. Ez utóbbira példa a baktérium Bacillus subtilis, amelyben MreB fehérjék, Mbl ( Mújra B-l ike) és MreBH ( MreB h omolog).
A genomokban E. coliÉs B. subtilis az MreB szintéziséért felelős gén ugyanabban az operonban található, mint az MreC és MreD fehérjék génjei. Az ezen operon expresszióját elnyomó mutációk csökkent életképességű gömbsejtek kialakulásához vezetnek.
Az MreB fehérje alegységei szálakat alkotnak, amelyek egy pálcika alakú baktériumsejt köré tekerednek. A citoplazma membrán belső felületén helyezkednek el. Az MreB által alkotott filamentumok dinamikusak, folyamatosan polimerizálódnak és depolimerizálódnak. Közvetlenül a sejtosztódás előtt az MreB azon a területen koncentrálódik, ahol a szűkület kialakul. Úgy gondolják, hogy az MreB funkciója a murein, egy sejtfal polimer szintézisének koordinálása is.
Az MreB homológok szintéziséért felelős gének csak rúd alakú baktériumokban voltak megtalálhatók, coccusokban nem.
A ParM fehérje alacsony kópiaszámú plazmidokat tartalmazó sejtekben van jelen. Feladata a plazmidok hígítása a sejt pólusai mentén. Ugyanakkor a fehérje alegységei a pálcika alakú sejt nagy tengelye mentén kifeszített filamentumokat alkotnak.
Az izzószál szerkezetében kettős spirál. A ParM által alkotott filamentumok növekedése mindkét végén lehetséges, ellentétben az aktin filamentumokkal, amelyek csak a ±póluson nőnek.
A MamK egy aktinszerű fehérje Magnetospirillum magneticum felelős a magnetoszómák helyes elhelyezkedéséért. A magnetoszómák a vasrészecskéket körülvevő citoplazma membrán invaginációi. A MamK filamentum vezetőként működik, amely mentén a magnetoszómák egymás után helyezkednek el. A MamK fehérje hiányában a magnetoszómák véletlenszerűen oszlanak el a sejtfelszínen.
Jelenleg két tubulin homológot találtak prokariótákban: FtsZ és BtubA/B. Az eukarióta tubulinhoz hasonlóan ezek a fehérjék GTPáz aktivitással rendelkeznek.
Az FtsZ fehérje rendkívül fontos a baktériumsejtek osztódásában, szinte minden eubaktériumban és archaeában megtalálható. Ezen túlmenően ennek a fehérjének homológjait találták eukarióta plasztidokban, ami szimbiotikus eredetük újabb megerősítése.
Az FtsZ az úgynevezett Z-gyűrűt alkotja, amely további sejtosztódási fehérjék vázaként működik. Ezek együttesen alkotják a szűkület (septa) kialakulásáért felelős szerkezetet.
A széles körben elterjedt FtsZ-vel ellentétben ezek a fehérjék csak a nemzetséghez tartozó baktériumokban találhatók meg Prosthecobacter. Szerkezetükben közelebb állnak a tubulinhoz, mint az FtsZ.
A fehérjét a sejtekben találták meg Caulobacter crescentus. Feladata sejteket adni C. crescentus vibrio formák. A sejtkreszcentin gén expressziójának hiányában C. crescentus bot formáját ölti. Érdekes módon a kettős mutánsok, a crescentin - és az MreB - sejtjei gömb alakúak.
Ezeknek a fehérjéknek nincs homológja az eukarióták között.
A MinD felelős a baktériumok és plasztidok osztódási helyének helyzetéért. A ParA részt vesz a DNS leánysejtekre való osztódásában.
|
||||||||||||||||||||||
Anna Pavlovna szalonja fokozatosan megtelt. Megérkezett Szentpétervár legmagasabb nemessége, a legheterogénebb korú és jellemű emberek, de ugyanazok a társadalomban, amelyben mindenki élt; megérkezett Vaszilij herceg lánya, a gyönyörű Heléna, aki behívta apját, hogy menjen el vele a követ lakomájára. Ciprusban és báli ruhában volt. La femme la plus seduisante de Petersbourg [a legbájosabb nő Szentpéterváron] néven is ismert a fiatal, kis Bolkonszkaja hercegnő, aki tavaly télen férjhez ment, és most nem a terhesség miatt ment ki a nagyvilágba, hanem elment. kis estéken is megérkezett. Hippolit herceg, Vaszilij herceg fia megérkezett Mortemarral, akit bemutatott; Morio abbé és még sokan mások is eljöttek.
- Még nem láttad? vagy: - nem ismered a ma tante-t [a nénikémmel]? - mondta Anna Pavlovna a látogató vendégeknek, és nagyon komolyan odavezette őket egy kis öregasszonyhoz, aki magasan meghajolt, aki egy másik szobából lebegett, amint elkezdtek érkezni a vendégek, a nevén szólította őket, lassan elfordította a tekintetét vendége volt ma tante [nagynéninek], majd elment.
Valamennyi vendég elvégezte a senkinek ismeretlen, érdektelen és szükségtelen néni köszöntésének szertartását. Anna Pavlovna szomorú, ünnepélyes részvéttel követte üdvözletüket, hallgatólagosan helyeselve őket. Ma tante mindenkivel egyformán beszélt az egészségéről, az ő egészségéről és Őfelsége egészségéről, amely ma, hála Istennek, jobb volt. Mindazok, akik tisztességből sietség nélkül, az elvégzett nehéz kötelességtől megkönnyebbülten közeledtek, eltávolodtak az öregasszonytól, hogy ne menjenek fel hozzá egész este.
A fiatal Bolkonszkaja hercegnő munkával érkezett egy hímzett arany bársonytáskában. Csinos, enyhén megfeketedett bajuszú, felső ajka fogaiban rövid volt, de annál szebben kinyílt, és néha még szebben kinyúlt, és ráesett az alsóra. Mint mindig az igen vonzó nőknél, rövid ajkak és félig nyitott szája a különlegessége, a saját szépsége volt. Mindenkinek jó móka volt nézni ezt a csinos, egészséggel és elevenséggel teli leendő anyukát, aki olyan könnyen tűrte helyzetét. Az öregeknek és a ránéző unatkozó, komor fiataloknak úgy tűnt, ők maguk is olyanná válnak, mint ő, miután egy ideig vele voltak és beszélgettek. Bárki, aki beszélt vele, és minden szavánál látta ragyogó mosolyát és folyamatosan láthatóan csillogó fehér fogait, azt gondolta, hogy ma különösen kedves. És mindenki ezt gondolta.
A kis hercegnő kacsázva, kis gyors léptekkel, munkatáskával a karján megkerülte az asztalt, és vidáman megigazgatta a ruháját, leült a kanapéra, az ezüst szamovár mellé, mintha minden, amit csinált volna, egy rész de plaisir [szórakozás ] neki és a körülötte lévőknek.
- J "ai apporte mon ouvrage [megragadtam az állást]" - mondta, és kibontotta a táskáját, és együtt szólított meg mindenkit.
– Nézd, Annette, ne me jouez pas un mauvais tour – fordult a háziasszonyhoz. - Vous m "avez ecrit, que c" etait une toute petite soiree; voyez, comme je suis attifee. [Ne játssz velem rossz tréfát; azt írtad nekem, hogy nagyon kicsi estéd volt. Nézd, milyen rosszul vagyok felöltözve.]
És széttárta a kezét, hogy megmutasson neki egy elegáns szürke ruhát csipkében, széles szalaggal övezve kicsit a melle alatt.
- Soyez tranquille, Lise, vous serez toujours la plus jolie [Légy nyugodt, te leszel a legjobb] - válaszolta Anna Pavlovna.
- Vous savez, mon mari m "abandonne" - folytatta ugyanazon a hangon, a tábornokra utalva - "il va se faire tuer. Dites moi, pourquoi cette vilaine guerre, [Tudod, a férjem elhagy. Mondd, miért ez a csúnya háború.] - mondta Vaszilij hercegnek, és válaszra sem várva Vaszilij herceg lányához, a gyönyörű Helenához fordult.
- Quelle delicieuse personne, que cette petite princesse! [Milyen bájos ember ez a kis hercegnő!] - mondta Vaszilij herceg csendesen Anna Pavlovnának.
Nem sokkal a kis hercegnő után egy masszív, termetes fiatalember lépett be, nyírt fejjel, szemüveges, korabeli divatos világos nadrágos, magas sallangú, barna frakkos fiatalember. Ez a kövér fiatalember Katalin híres nemesének, Bezukhoi grófnak volt törvénytelen fia, aki most Moszkvában haldoklott. Még nem szolgált sehol, most érkezett külföldről, ahol nevelkedett, és először került a társadalomba. Anna Pavlovna meghajlással üdvözölte, amely szalonjában a legalacsonyabb hierarchia embereié volt. Ám e gyengébb üdvözlés ellenére Pierre belépése láttán Anna Pavlovna szorongást és félelmet tanúsított, hasonlóan ahhoz, ami egy helyhez képest túl hatalmas és szokatlan dolog láttán nyilvánul meg. Noha Pierre valóban valamivel nagyobb volt, mint a többi férfi a szobában, de ez a félelem csak ahhoz az intelligens és egyben félénk, figyelmes és természetes tekintethez kapcsolódott, amely megkülönböztette őt ebben a nappaliban mindenkitől.
- ez egy fonalas szerkezetek rendszere, ami a legfontosabb, hogy azonos osztályba tartozó fehérjék rendezett polimerei, amelyek a baktériumok és az archaea sejtjeiben jelen vannak. A bakteriális citoszkeleton összes vizsgált (2006-tól) fehérjéje képes hosszú filamentumokká önszerveződni. in vitro.
A prokarióták citoszkeletonját először az 1990-es évek elején fedezték fel, amikor kiderült, hogy szinte minden baktérium és a legtöbb archaea tartalmazza az FtsZ fehérjét, amely tubulin homológ, és a sejtosztódás során gyűrűképző filamentumokká (Z-gyűrű) polimerizálódhat. Később az aktin prokarióta homológjait is felfedezték. Ezek a felfedezések megváltoztatták azt az elképzelést, hogy a citoszkeleton hiánya a legfontosabb oka a prokarióták kisebb méretének és egyszerűbb szerveződésének az eukariótákhoz képest. Másrészt ma már azt feltételezik, hogy a baktériumok és az archaeák viszonylagos egyszerűsége összefügg a motoros fehérjék jelenlétével (legalábbis eddig még nem fedezték fel), amelyek a citoszkeleton filamentumai mentén „sétálnak” és biztosítanak különböző struktúrák szállítására, valamint az egész sejt mozgására.
Az aktin és tubulin homológjainak jelenléte prokariótákban arra utal, hogy a nukleotidkötő fehérjék e két osztálya, amelyek dogwie filamentumokat képezhetnek, meglehetősen régen, még az eukarióták megjelenése előtt az evolúció során keletkeztek. A nukleáris és nem nukleáris szervezetek azonban eltérően használják őket, például a tubulin homológ FtsZ részt vesz a bakteriális citokinézisben, míg az aktin filamentumok látják el ezt a funkciót az eukariótákban, éppen ellenkezőleg, az aktin homológok részt vesznek a DNS-molekulák közötti különbségben az osztódás során. baktériumok és mikrotubulusok eukariótákban, tubulinnal, amely az osztódás orsóját alkotja. Ezenkívül legalább egy fehérjeosztályt találtak prokariótákban, amelyek a közbenső filamentum fehérjék homológjainak tekinthetők, és a citoszkeletális fehérjék egy osztálya - Walker A típusú ATPáz (WACA - MinD és PraA), amelyek nem mutatnak megfelelőséget az eukariótákban.
2001-ben Jones Jones)és spivrobintniki megállapította, hogy a baktérium Bacillus subtilis aktin homológ fehérjék vannak jelen, amelyek hosszú spirális szerkezeteket alkotnak. Ez a felfedezés intenzív kutatásfejlesztést indított el a prokarióták citoszkeletonja területén, melynek eredményeként számos más aktin homológot fedeztek fel. Mindezeket a fehérjéket egy aktin ATPáz domén jelenléte jellemzi. Legtöbbjük, mint az eukarióták aktinja, a citoszkeleton része, de némelyik más funkciót is ellát, például a sejtosztódásban részt vevő FtsA, a DnaK chaperon és a hexokináz. A bakteriális aktin homológok hasonló térszerkezetűek, de többnyire elég erősen különböznek aminosav-szekvenciában (5-10%-os azonosság). Ezen túlmenően ezek a fehérjék kiváló tulajdonságokkal rendelkeznek a polimerizáció dinamikája és az általuk képzett filamentumok tulajdonságai tekintetében. Nyilvánvaló, hogy az eukariótáktól eltérően, amelyek ugyanazt az aktint használják különféle sejtszükségletekhez, a baktériumoknak számos ilyen fehérje változata van, amelyek mindegyike külön funkció ellátására specializálódott.
mreb (angol) M te irányítod a B klasztert)és homológjai - a fehérjék gyakoriak a pálcika vagy spirál alakú baktériumok között, és hiányoznak a coccusokból. Néhány baktérium pl Escherichia coliÉs Caulobacter crescentus, csak az MreB fehérje gént tartalmazza, míg mások, különösen bacillus subtilis, ezen kívül homológjainak génjei Mbl (eng. M re B — mint) és MreBH MreB h omolog). Ezek a fehérjék biztosítják a sejt rúd alakú formájának megőrzését, polaritását, valamint a bakteriális DNS kópiák közötti különbségeket az osztódás során.
in vivo Az MreB fehérje és homológjai a baktériumsejt mentén elhelyezkedő hosszú spirális filamenteket alkotnak, amelyek erős és meglehetősen rugalmas kötegekké kombinálhatók. Az ilyen szálak dinamikus szerkezetek, felezési idejük általában nem haladja meg a néhány percet. Ezen kívül egyes fajoknál különösen C.crescentusÉs Rhodobacter sphaeroides Az MreB filamentumok a sejtciklus során megváltoztatják elhelyezkedésüket: osztódáskor a sejt központi részében koncentrálódnak és gyűrűt alkotnak. Mivel azonban az mreB deléciós mutánsok nem veszítik el képességüket a citokinézisre, úgy tűnik, hogy az MreB fehérje nem szükséges ehhez a folyamathoz.
Amint azt a bakteriális fehérjéken végzett kísérletek mutatják Thermotoga maritima Az MreB monomer egységek önszerveződésre képesek in vitro hosszú lineáris filamentumokká, amelyek két párhuzamosan elhelyezkedő protofilamentumból állnak. Tehát az MreB filamentumok szerkezete szerint F-aktinban különböznek egymástól, amelyet két spirálisan egymás köré csavart lánc alkot. Az MreB polimerizációjához ATP jelenléte szükséges a közegben, de ugyanolyan jól megy végbe GTP jelenlétében (ellentétben az aktinnal, amely csak ATP jelenlétében polimerizálódik). Ennek oka az a tény, hogy a polimerbe csak nukleotid-trifoszfáttal társított formában kerülnek be új alegységek, később megtörténik a megkötött ATP vagy GTP hidrolízise ADP-vé, illetve GDP-vé.
Az MreB filamentumok és homológ fehérjék egyik fő funkciója a baktériumsejt rúd- vagy spirális alakjának megőrzése. Az ezen fehérjék expresszióját megzavaró mutációk a baktériumok kifejezett alakváltozását eredményezik (általában lekerekített sejtekké, vagy Mbl esetében szabálytalan alakú sejtekké alakulnak). Az MreB filamentumok azonban NEM szolgálnak közvetlenül a sejtalak vázaként, hanem spirálisan elrendezve, a sejtfal peptidoglikánját szintetizáló enzimek kötődési helyei. Így szabályozzák az új elemek baktériumhéjra való lerakódásának természetét, ami valójában a meghatározó tényező az állandó forma fenntartásában. Hasonlóképpen, a növényi sejt mikrotubulusai befolyásolják annak alakját azáltal, hogy cellulózmolekulák zárványait a sejtfalba irányítják. Sok baktériumban (beleértve E. coliÉs B.subtilis) gén mreB az operon része, amely géneket is tartalmaz mreCÉs mreD. Ez az operon a peptidoglikán bioszintéziséhez szükséges gének nagy csoportjába tartozik. Gén termékek mreCÉs mreD Gram-negatív baktériumok belső membránjának fehérjéi, kölcsönhatásba lépnek az MreB fehérjével, és részt vesznek a murein bioszintézisben részt vevő enzimekkel, mint például a murein transzpeptidáz PBP2 komplexének megszervezésében. Ez a komplex tartalmazza a RodZ és RodA transzmembrán fehérjéket is.
Az MreB filamentumok részt vesznek a sejtpolaritás bizonyos aspektusainak meghatározásában is, különösen bizonyos fehérjék egyik vagy mindkét pólusán, például a kemotaxisért, mozgékonyságért, szekrécióért és virulenciáért felelősek koncentrációjában.
Az MreB és homológjainak másik funkciója a bakteriális kromoszóma másolatai közötti különbségben való részvétel az osztódás során. A mutánsok között, amelyekben ez a fehérje hiányzik, olyan sejteket találtak, amelyekben számos nukleoid található a citoplazmában, valamint olyan sejteket, amelyeknek nem volt kromoszómájuk. Az MreB fehérjék bakteriális DNS-hez való kapcsolódási helye az oriC pont, a kapcsolódás közvetlenül vagy más fehérjék részvételével történik. Osztva a citoszkeleton filamentumok különbségeket adnak két DNS-másolat oriC pontjaiban a sejt ellentétes végén, ennek a folyamatnak a mechanizmusa még nem tisztázott (2006). Az sem ismert, hogy a kromoszóma szegregáció hogyan megy végbe olyan coccusokban, amelyekből hiányzik a gén mreBés homológjai.
Sok alacsony kópiaszámú (~ 1-5 kópia) bakteriális plazmid speciális rendszerrel rendelkezik, amely biztosítja a replikáció utáni különbségeiket. Ezek a mechanizmusok szükségesek ahhoz, hogy az egyes leánysejtek legalább egy plazmid DNS-molekulát kapjanak az elválasztás után. Háromféle rendszer létezik, amelyek különbségeket biztosítanak az alacsony kópiaszámú plazmidokban, amelyek mindegyike különböző motorfehérjéket használ (I-es típusú - Walker A-típusú ATPázok vagy ParA-szerű fehérjék, II-es típusú - tubulin homológok vagy TubZ-alakú fehérjék, III-as típus - aktin homológok vagy ParM alakú fehérjék). Protein ParM (angol nyelvből. Par pozicionáló motor) először a plazmidok tanulmányozása során fedezték fel R1 E. coli. Ez a plazmid DNS szegregációs rendszer ma már jobban érthető. Hasonló rendszert találtak más plazmidokban is, különösen azokban, amelyek felelősek a multidrog rezisztencia terjedéséért. multidrog rezisztencia).
Mint a citoszkeleton minden eleme, a ParM filamentumok is monomer fehérje alegységekből állnak. Ezek az alegységek polimerizációra képesek in vitro ATP vagy GTP jelenlétében. Az így létrejövő filamentumok két egymás köré csavart protofilamentumból állnak (szerkezete hasonló az F-aktinhoz). Az élő sejtekben a ParM monomerek hosszú, el nem ágazó szálakat képeznek, amelyek a baktérium tengelye mentén helyezkednek el. Az aktinnal és az MreB-vel és analógjaival ellentétben a ParM nem alkot kötegeket.
A ParM monomerek polimerizációja és disszociációja az ATP addíciójától és hidrolízisétől függ. Az új alegységek ATP-hez kötött formában épülnek be a filamentumokba, és a kötődés a filamentumok mindkét végén megtörténhet. Az új ParM-ATP alegység felvételével egyidejűleg az ATP hidrolízise megy végbe az utoljára kapcsolódó fehérjemolekulában. Így az egész filamentum ParM-ADP fehérjékből áll, és csak a végein találhatók ParM-ATP alegységek, amelyek a teljes szerkezetet "KEPU" stabilizálják.
A megfelelő plazmid hiányában a ParM filamentumok tovább polimerizálódnak, amíg el nem érnek egy bizonyos kritikus hosszúságot. Ezt követően nagyon gyorsan disszociálni kezdenek, és ennek a folyamatnak a sebessége körülbelül 100-szor nagyobb, mint az F-aktiné, vagyis megfigyelhető az úgynevezett dinamikus instabilitás, amely szerint ezek az elemek inkább eukarióta mikrotubulusokhoz hasonlítanak. .
Gén parM belép a lokuszba par R1 plazmidot, ezen kívül egy szakaszt is tartalmaz parC(angolról. C entromere), amely a centromerhez hasonló szerepet játszik az eukarióta kromoszómákban, valamint a gén parr, amelynek terméke a ParR (angol. Represszor) csatlakozik a szekcióhoz parCés autoregulálja a lókusz transzkripcióját par,és adapterként is szolgál a ParM fehérje rögzítéséhez.
Az R1 plazmid replikációja után mindkét kópiájára a régióban parC a ParR fehérje kapcsolódik. Ebben az állapotban képes megkötni és stabilizálni a ParM filamentumokat, amelyek folyamatosan össze- és szétszerelődnek a citoplazmában. Ezt követően a ParM polimer szálak elkezdenek rágni, és mindkét végén új monomerek tapadnak. Ezt a folyamatot az ATP hidrolízise kíséri. A filamentumok megnyúlása miatt a széleihez kapcsolódó két plazmid különböző irányba hasad, amíg el nem érik a sejt pólusait. Ezt követi a ParM polimer disszociációja.
Az aktin másik prokarióta homológja, a MamK részt vesz a magnetoszóma membránok szerveződésében. A magnetoszómák a nemzetséghez tartozó baktériumok membránhoz kötött organellumai MagnetospirillumÉs Magnetococcus, magnetit kristályokat tartalmaznak, és segítik a baktériumot a geomágneses térben való navigálásban. A sejtben a magnetoszómák sorba rendeződnek, aminek következtében mágnestűként működhetnek. Ezt az elrendezést a MamK fehérje filamentumai biztosítják, amelyekhez ezek a hártyás vezikulák kapcsolódnak.
A legtöbb prokarióta a mikrotubulusokat alkotó eukarióta fehérje tubulin homológjaival is rendelkezik. E homológok közül a legjobban tanulmányozott a citokinézisben szerepet játszó FtsZ blilock. A tubulin és az FtsZ aminosavszekvenciájában meglehetősen csekély azonosságot mutat, csak a GTPáz domén konzervált, de térszerkezetükben hasonlóak. Ezenkívül a baktériumok és archaea egyes képviselőiben a tubulin más homológjait is találták: például BtubA / BtubB Prosthebacter dejoneii, valamint TubZ és RepX, amelyeket a nemzetségbe tartozó baktériumok plazmid génjei kódolnak bacilus.
FtsZ (FtsZ) F panaszkodó hőmérséklet- s enyhe mutáns Z) az egyik első prokariótákban azonosított citoszkeletális fehérje. Szinte az összes vizsgált baktérium és archaea sejtjében, valamint a prokariótákból származó eukarióta organellumokban, különösen a plasztidokban megtalálható. Ez a fehérje részt vesz a Z-gyűrű kialakításában, citokinézist biztosít a sejtosztódás során. Ebben a folyamatban az FtsZ mellett számos segédfehérje is részt vesz, különösen azok, amelyek a baktériumsejtfal szintézisében vesznek részt.
FtsZ monomerek keletkeznek in vitro protofilamentumok, amelyek ezen fehérjék egy sorából állnak. A protofilamentumok NEM állnak össze a mikrotubulusokhoz hasonló szerkezetekké, bár néha kötegeket vagy lapokat képeznek. Az FtsZ aktív GTP-kötött formában polimerizálódik, azonban a tubulinnal ellentétben ez a fehérje általában nem hidrolizálja a GTP-t a protofilamentumba való beépülése után. Így a mikrotubulus protofilamentumokkal szemben, amelyek szinte teljes egészében GDP-tubulinból állnak, és csak a végén GTP-tubulin sapkával rendelkeznek, a GTP-hez kötött és a GDP-hez kötött alegységek aránya az FtsZ protofilamentumokban 80:20.
Az FtsZ protofilamentumokban bizonyos körülmények között GTP hidrolízis léphet fel, ilyenkor az alakjuk elsősorban egyenesből ívessé változik, és a polimer destabilizálódik, aminek következtében monomerekre bomlik. Az FtsZ protofilamentumok dinamikus struktúrák, folyamatosan cserélik az alegységeket szabad monomerek készletével.
A sejtben található FtsZ fehérje egy része a Z-gyűrű kialakításában vesz részt, míg a többi monomer formában, vagy rövid filamentumok formájában a citoplazmában található. Amint azt a fluoreszcens mikroszkóppal kimutatták (jelölt antitestek vagy GFP-hez fuzionált FtsZ használatával), a Z-gyűrű jól látható a legtöbb sejt közepén. A sejtosztódás során összehúzódik, ezáltal lehetővé teszi a citokinézist. Az anyasejtben a Z-gyűrű redukciójával egyidejűleg az FtsZ polimerizálódni kezd a leánysejtek közepén.
A Z-gyűrű nem egy, a protofilamentumba zárt FtsZ-ből áll, amint azt számos tanulmány kimutatta, a Z-gyűrűben lévő FtsZ monomerek mennyisége elegendő ahhoz, hogy körülbelül 2,5 fordulatot tegyen a cella belső átmérője körül. Mivel az egyes FtsZ protofilamentumok jóval rövidebbek, mint a sejt kerülete, ezért javasoltam a Z-gyűrű szerkezetének modelljét, amely szerint az nagyszámú, rövid átfedő profilból áll. Ezt a modellt elektronikus kriotomográfiával nyert adatok igazolták. Vannak azonban alternatív modellek is a Z-gyűrű szerkezetére, amelyek közül az egyik azt feltételezi, hogy az FtsZ protofilamentumok végponttól végig kölcsönhatásba lépnek, és egy folytonos hélixet alkotnak.
A citokinézis lehetővé tételéhez a Z-gyűrűt valahogyan hozzá kell kötni a plazmamembránhoz. Ezt a szerepet a legtöbb baktériumban az FtsA piintegrális fehérje és a transzmembrán ZipA fehérje tölti be, amelyek citoplazmatikus doménjei az FtsZ-hez kapcsolódnak.
A Z-gyűrű összehúzódásának mechanizmusa a citokinézis során még mindig nem tisztázott. Számos hipotézis létezett, amelyeket fent leírtunk:
Számos baktérium genomjának szekvenálása feltárt néhány tubulinszerű fehérjét, amelyek különböznek az FtsZ-től. Főleg a baktériumokban Prosthebacter dejoneii két BtubA és BtubB fehérjét találtak. B aktuális kád ulin), amelyek az α illetve β tubulin homológjai. A GTP jelenlétében történő polimerizáció során heterodimert képeznek, akárcsak az α és β tubulin. Ezeknek a fehérjéknek a funkciója jelenleg nem ismert.
Érdekes módon ezek a fehérjék aminosav-szekvenciájában sokkal közelebb állnak az eukarióta tubulinokhoz, mint a prokarióta homológjukhoz, az FtsZ-hez. Úgy tartják, hogy a baktérium P. dejoneii eukariótákból horizontális átvitel eredményeként kapta meg ezeknek a fehérjéknek a génjeit.
A tubulin homológok egy másik osztályát a nemzetségbe tartozó baktériumok nagy plazmidjaiban találták meg bacilus, zokema:
Mindkét fehérje képes hosszú filamentumokat képezni a GTP jelenlétében történő polimerizáció eredményeként, és szükséges a megfelelő plazmid sejtben való stabil fenntartásához. Részt vehetnek a plazmid kópia szegregációjában, a plazmid replikációjában vagy mindkettőben.
A kreszcentin egy köztes filamentum fehérje, amely baktériumokban található Caulobacter crescentusés ebbe a nemzetségbe tartozó más baktériumok. Ez a fehérje hosszú, ívelt, fonalas szerkezetet tart fenn, amely a como-szerű baktérium belső széle mentén fut, és biztosítja ennek az alaknak a megőrzését. Kreszcentin hiányában a baktériumok lepényszerűvé válnak, de nem veszítik el vitalitásukat.A Crescentin 25%-os azonosságot és 40%-os homológiát mutat aminosav-szekvenciában az eukarióta köztes filamentumfehérjékkel, valamint a fehérjedomének hasonló szerveződése – különösen , egy központi kettős hélix domén jelenléte (eng. tekercselt tekercs). A crescentin monomerek polimerizációja, mint az eukarióta intermedier filamentum fehérjék esetében, nukleotidok nélkül megy végbe. Kíváncsi vagyok, mit formáljak C.crescentus a crescentin mellett az MreB aktin homológra is szükség van, ennek hiányában a sejtek a kreszcentin jelenléte ellenére gömb alakúvá válnak.
Az eukarióta aktin, a tubulin és a köztes filamentum fehérjék homológjain kívül olyan citoszkeletális komponenseket is találtak olyan baktériumokban, amelyeknek nincs analógja a sejtmagban. Ezek különösen a WACA fehérjék (eng. Walker A citoszkeletális ATPáz- Walker A típusú citoszkeletális ATPáz), amelyek az ATP-ázok funkcionálisan heterogén családjába tartoznak, szerkezetükben konzervatív abnormális Walker A doménnel és ATP jelenlétében dimerizálódnak.
A WACA fehérjék ATP-kötött formában polimereket képezhetnek bizonyos felületeken, például a sejtmembránon, és a citoszkeleton elemeinek tekinthetők. Ebbe az osztályba tartozik a MinD fehérje, amely az elválasztás során a citokinézis helyének meghatározásában vesz részt, valamint a ParA, Soj, valamint SopA és ParF fehérjék, amelyek a plazmid és a bakteriális kromoszóma kópiák különbségeit (szegregációját) biztosítják. Bár eltérő funkciójuk van, ezek a fehérjék nagyon hasonló térszerkezettel és magas szintű aminosavszekvencia-homológiával rendelkeznek. Minden WACA képes ATP hidrolízisre, katalitikus aktivitását az aktiváló fehérjékkel való kölcsönhatás szabályozza: MinD esetében ez a MinE fehérje, ParA esetében pedig a DNS-kötő ParB fehérje. Ezenkívül ezt a fehérjecsaládot egyesíti az a tény, hogy mindegyik mögött dinamikus viselkedés figyelhető meg. in vivo: e fehérjék polimerizált formái bizonyos sejtrégiók között oszcillálnak. Például a MinD-k először a cella egyik pólusán polimerizálódnak, majd a másikon, egy ilyen ciklus időtartama 40-50 másodperc. A ParA és Soj fehérjék döntően két nukleoid között oszcillálnak a hasadás előtt, és „ugrási” időintervallumuk kevésbé szabályos (több perctől egy óráig).
Az oszcillációs mechanizmus jobban megérthető a MinCDE rendszer példáján, amely magában foglalja a WACA MinD-t. Ez a rendszer szükséges a sejt számára ahhoz, hogy a Z-gyűrűt pontosan elhelyezze a központi részbe a citokinézis helyes áthaladásához. Három fehérjét tartalmaz:
BAN BEN E. coli ez a rendszer a következőképpen működik: ATP molekula hozzáadása után a MinD polimerizálódik a plazmamembránon, spirálokat képezve. Ebben az aktivált formában a MinC fehérjéhez kötődik, amely gátolja a Z-gyűrű kialakulását az adott helyen. A MinD-ATP kölcsönhatásba léphet a MinE-vel is, ami stimulálja az ATP hidrolízist, ami után az inaktivált MinD leválik a membránról és máshol disszociálhat. Főleg a sejt ellenpólusára bomlik fel, ahol nincs MinE fehérje, ott kezdődik meg az új komplex polimerizációja, ami addig tart, amíg a régi depolimerizációja be nem fejeződik. És amikor elkezd véget érni, a MinE fehérje felszabadul, és elkezdi „megsemmisíteni” az újonnan képződött MinD / MinC komplexet. Így ez a komplexum 40-50 perces periodikusan "ugrik" egyik pólusról a másikra, és nem csak a központi területet érinti, ahol a Z-gyűrű kialakulása történik, mivel ott semmi sem nyomja el.
Bár a MinD nagyon konzervált fehérje a prokarióták között, különböző fajokban eltérően működik, pl. B. subtilis nem történik oszcilláció: A MinD egy másik DivIVA fehérje segítségével tartósan a sejtpólusokhoz kapcsolódik. Emellett a baktériumoknak „tartalék” mechanizmusai vannak a citokinézis térbeli szabályozására, amelyek még a MinCDE hiányában is működnek, ilyen például a „nukleoid elkerülő” mechanizmus: a Z-gyűrű kialakulását a nukleoid közelében elnyomják.
Egyes baktériumokban mind a MinCDE rendszer, mind a „nukleoid elkerülő” mechanizmus teljesen hiányzik, például C.crescentus a citokinézis helyét a MipZ fehérje határozza meg (hasonlóan a ParA-hoz). Ez a fehérje az ori pont közelében polimerizálódik, és gátolja a Z-gyűrű kialakulását is.
eukarióta citoszkeleton. Az aktin mikrofilamentumok vörösre, a mikrotubulusok zöldre, a sejtmagok kékre festődnek.
Citoszkeleton- ez egy sejtváz vagy váz, amely egy élő sejt citoplazmájában található. Az eukarióták és prokarióták minden sejtjében jelen van. Ez egy dinamikus, változó struktúra, melynek funkciói közé tartozik a sejt alakjának megtartása és hozzáigazítása a külső hatásokhoz, exo- és endocitózis, a sejt egészének mozgásának biztosítása, aktív intracelluláris transzport és sejtosztódás.
Keratin közbenső filamentumok egy sejtben.
A citoszkeleton fehérjékből áll. A citoszkeletonban több fő rendszert különböztetnek meg, amelyeket vagy az elektronmikroszkópos vizsgálatok során látható főbb szerkezeti elemek (mikrofilamentumok, intermedier filamentumok, mikrotubulusok), vagy az ezeket felépítő fő fehérjék (aktin-miozin rendszer, keratinok, tubulin) alapján neveznek el. -dynein rendszer).
Az eukarióta sejtek háromféle úgynevezett filamentumot tartalmaznak. Ezek szupramolekuláris, kiterjesztett szerkezetek, amelyek azonos típusú, polimerekhez hasonló fehérjékből állnak. A különbség abban rejlik, hogy a polimerekben a monomerek közötti kötés kovalens, míg a filamentumokban az alkotóegységek kötése gyenge nem kovalens kölcsönhatás miatt jön létre.
A körülbelül 7 nm átmérőjű mikrofilamentumok két spirális aktinmonomer lánc. Főleg a sejt külső membránján koncentrálódnak, mivel a sejt alakjáért felelősek, és képesek a sejtfelszínen kiemelkedéseket (pszeudopodiákat és mikrobolyhokat) kialakítani. Részt vesznek az intercelluláris interakcióban (tapadó kontaktusok kialakításában), a jelátvitelben és a miozinnal együtt az izomösszehúzódásban is. A citoplazmatikus miozinok segítségével mikrofilamentumok mentén vezikuláris transzport valósítható meg.
A közbenső szálak átmérője 8-11 nanométer. Különféle alegységekből állnak, és a citoszkeleton legkevésbé dinamikus részét képezik.
Diagram, amely a citoplazmát mutatja komponenseivel együtt (vagy sejtszervecskék) egy tipikus állati sejtben. Sejtszervecskék:
(1) Nucleolus
(2) Mag
(3) riboszóma (kis pontok)
(4) Hólyagos
(5) durva endoplazmatikus retikulum (ER)
(6) Golgi-készülék
(7) Citoszkeleton
(8) Sima endoplazmatikus retikulum
(9) Mitokondriumok
(10) Vacuole
(11) Citoplazma
(12) Lizoszóma
(13) Centriole és Centrosome
A mikrotubulusok körülbelül 25 nm átmérőjű üreges hengerek, amelyek fala 13 protofilamentumból áll, amelyek mindegyike tubulin fehérje dimer lineáris polimerje. A dimer két alegységből áll - a tubulin alfa és béta formáiból. A mikrotubulusok rendkívül dinamikus struktúrák, amelyek a polimerizáció során GTP-t fogyasztanak. Kulcsszerepet játszanak az intracelluláris transzportban ("sínként" szolgálnak, amelyen a molekuláris motorok - kinezin és dynein mozognak), a mitózis és a meiózis során az undylipodium axoneme és az osztódási orsó alapját képezik.
Sokáig azt hitték, hogy csak az eukariótáknak van citoszkeletonja. Azonban Jones és munkatársai 2001-es tanulmánya óta. (PMID: 11290328), amely leírja a bakteriális aktin homológok szerepét a sejtekben Bacillus subtilis, megkezdődött a bakteriális citoszkeleton elemeinek aktív tanulmányozásának időszaka. A mai napig bakteriális homológokat találtak mindhárom típusú eukarióta citoszkeleton elemhez - a tubulinhoz, az aktinhoz és a köztes filamentumokhoz. Azt is megállapították, hogy a bakteriális citoszkeleton fehérjék legalább egy csoportja, a MinD/ParA, nem rendelkezik eukarióta analógokkal.
A citoszkeleton leginkább vizsgált aktinszerű komponensei az MreB, a ParM és a MamK.
Az MreB fehérjék és homológjai a bakteriális citoszkeleton aktinszerű komponensei, amelyek fontos szerepet játszanak a sejtforma fenntartásában, a kromoszóma szegregációban és a membránszerkezetek szerveződésében. Egyes baktériumfajták, mint pl Escherichia coli, csak egy MreB fehérjét tartalmaznak, míg másokban 2 vagy több MreB-szerű fehérje található. Ez utóbbira példa a baktérium Bacillus subtilis, amelyben MreB fehérjék, Mbl ( Mújra B-l ike) és MreBH ( MreB h omolog).
A genomokban E. coliÉs B. subtilis az MreB szintéziséért felelős gén ugyanabban az operonban található, mint az MreC és MreD fehérjék génjei. Az ezen operon expresszióját elnyomó mutációk csökkent életképességű gömbsejtek kialakulásához vezetnek.
Az MreB fehérje alegységei szálakat alkotnak, amelyek egy pálcika alakú baktériumsejt köré tekerednek. A citoplazma membrán belső felületén helyezkednek el. Az MreB által alkotott filamentumok dinamikusak, folyamatosan polimerizálódnak és depolimerizálódnak. Közvetlenül a sejtosztódás előtt az MreB azon a területen koncentrálódik, ahol a szűkület kialakul. Úgy gondolják, hogy az MreB funkciója a murein, egy sejtfal polimer szintézisének koordinálása is.
Az MreB homológok szintéziséért felelős gének csak rúd alakú baktériumokban voltak megtalálhatók, coccusokban nem.
A ParM fehérje alacsony kópiaszámú plazmidokat tartalmazó sejtekben van jelen. Feladata a plazmidok hígítása a sejt pólusai mentén. Ugyanakkor a fehérje alegységei a pálcika alakú sejt nagy tengelye mentén kifeszített filamentumokat alkotnak.
Az izzószál szerkezetében kettős spirál. A ParM által alkotott filamentumok növekedése mindkét végén lehetséges, ellentétben az aktin filamentumokkal, amelyek csak a ±póluson nőnek.
A MamK egy aktinszerű fehérje Magnetospirillum magneticum felelős a magnetoszómák helyes elhelyezkedéséért. A magnetoszómák a vasrészecskéket körülvevő citoplazma membrán invaginációi. A MamK filamentum vezetőként működik, amely mentén a magnetoszómák egymás után helyezkednek el. A MamK fehérje hiányában a magnetoszómák véletlenszerűen oszlanak el a sejtfelszínen.
Jelenleg két tubulin homológot találtak prokariótákban: FtsZ és BtubA/B. Az eukarióta tubulinhoz hasonlóan ezek a fehérjék GTPáz aktivitással rendelkeznek.
Az FtsZ fehérje rendkívül fontos a baktériumsejtek osztódásában, szinte minden eubaktériumban és archaeában megtalálható. Ezen túlmenően ennek a fehérjének homológjait találták eukarióta plasztidokban, ami szimbiotikus eredetük újabb megerősítése.
Az FtsZ az úgynevezett Z-gyűrűt alkotja, amely további sejtosztódási fehérjék vázaként működik. Ezek együttesen alkotják a szűkület (septa) kialakulásáért felelős szerkezetet.
A széles körben elterjedt FtsZ-vel ellentétben ezek a fehérjék csak a nemzetséghez tartozó baktériumokban találhatók meg Prosthecobacter. Szerkezetükben közelebb állnak a tubulinhoz, mint az FtsZ.
A fehérjét a sejtekben találták meg Caulobacter crescentus. Feladata sejteket adni C. crescentus vibrio formák. A sejtkreszcentin gén expressziójának hiányában C. crescentus bot formáját ölti. Érdekes módon a kettős mutánsok, a crescentin - és az MreB - sejtjei gömb alakúak.
Ezeknek a fehérjéknek nincs homológja az eukarióták között.
A MinD felelős a baktériumok és plasztidok osztódási helyének helyzetéért. A ParA részt vesz a DNS leánysejtekre való osztódásában.

Tartalom: Biológiailag aktív adalékanyag, amely egy légyes bogárból (vagy légy...

Ekaterina MirimanovaSystem mínusz 60. Revolution System mínusz 60 Ekaterina Mirimanovával"Rendszer mínusz 60. Forradalom"...

Az elnehezülés és a puffadás mind a hétköznapi túlevés, mind a súlyosabb emésztőrendszeri problémák okai...

A második vércsoport, az Rh-negatív, sok évvel ezelőtt jelent meg, amikor az ember felhagyott a vadászattal, és ...

AZ ANOREXIA JELENSÉGÉNEK PSZICHOLÓGIAI VONATKOZÁSAI (KÍSÉRLETI TANULMÁNY) T. V. Tarasova, E. V. Arsentjeva V...

Tartalom Tekintettel arra, hogy a bőr ezen a területen vékony, hajlamosabb a különféle foltok megjelenésére. Vörösség alatt...

vseslav Szo, 2015.10.17. - 20:50 A Vasileostrovskaya állomás az egyik legrégebbi állomás...

Leggyakrabban a kecskegombát (Suillus bovinus) a gombászok mindennapi beszédében kecskének nevezik. Ez a cső alakú...

Minden embernek voltak legalább egyszer álmai életében. Sokan nem tulajdonítanak nekik jelentőséget, de néhány...

Daedalus (kráter). Átmérő: 93 km Mélység: 3 km (NASA fotó) A Hold ősidők óta vonzza az emberek figyelmét. A II...

Az Egyesült Államokban február harmadik hétfőjén ünneplik az elnök napját. A 70-es évekig az ünnepségeket arra időzítették, hogy...

Az utolsó hívás napján csak egy szót szeretnék mondani szeretett osztályfőnökömnek: „Köszönöm.” De mennyi az...

A görögdinnyét ízletes és egészséges bogyónak tartják, magvai különösen hasznosak. Kicsi, sötétbarna vagy...

„A beszéd fejlődése 6 évesnél idősebb gyermekeknél” A gyermek nem született beszéddel. Nem lehet határozott választ adni...

Ekaterina Mirimanova rendszer mínusz 60. Revolution System mínusz 60 Ekaterina Mirimanovával "Rendszer mínusz 60 ....
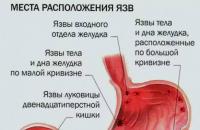
Az elnehezülés és a puffadás mind a hétköznapi túlevés, mind a komolyabb emésztési problémák okai.