Spanyol frontirányító két személyre – hogyan befolyásolja a libidót nőknél és férfiaknál
Tartalom Spanyol bogárból (vagy spanyol bogárból) nyert kivonat alapú étrend-kiegészítő...
A baktériumsejtnek nincs magja, a kromoszómák szabadon helyezkednek el a citoplazmában. Ezenkívül a baktériumsejtben hiányoznak a membránszervecskék: mitokondriumok, EPS, Golgi-készülék stb. A sejtmembrán külsejét sejtfal borítja.



A legtöbb baktérium passzívan mozog, víz- vagy légáramlatok segítségével. Csak néhányuknak van mozgásszervezete - flagella. A prokarióta flagellák nagyon egyszerű szerkezetűek, és flagellin fehérjéből állnak, amely 10-20 nm átmérőjű üreges hengert alkot. Becsavarják a táptalajt, előrehajtva a sejtet. Úgy tűnik, ez az egyetlen olyan szerkezet a természetben, amely a kerék elvét használja.
Alakjuk alapján a baktériumokat több csoportra osztják:
Cocci (kerek alakúak);
- bacillusok (rúd alakúak);
- spirilla (spirál alakú);
- vibrios (vessző alakú).
A légzés módja szerint a baktériumokat aerobokra (a legtöbb baktérium) és anaerobokra (tetanusz, botulizmus, gázgangréna kórokozói) osztják. Az előbbieknek oxigénre van szükségük a légzéshez; az utóbbiak számára az oxigén haszontalan, sőt mérgező.

A baktériumok protoplazmáját az eukariótákhoz hasonlóan plazmamembrán veszi körül. A membrán zsákszerű, tubuláris vagy lamellás invaginációi a légzési folyamatban részt vevő mezoszómákat, bakterioklorofillt és más pigmenteket tartalmaznak.
A prokarióták genetikai anyaga nem alkot magot, hanem közvetlenül a citoplazmában található. A bakteriális DNS egyetlen kör alakú molekula, amelyek mindegyike több ezer és millió nukleotidpárból áll. A baktériumsejt genomja sokkal egyszerűbb, mint a fejlettebb lények sejtjeié: a bakteriális DNS átlagosan több ezer gént tartalmaz.
A prokarióta sejtekben nincs endoplazmatikus retikulum, és a riboszómák szabadon lebegnek a citoplazmában. A prokariótáknak nincs mitokondriumuk; Feladatukat részben a sejtmembrán látja el.
A baktériumok mobilitását a flagellák biztosítják. A baktériumok körülbelül 20 percenkénti osztódással szaporodnak (kedvező körülmények között). A DNS replikálódik, és minden leánysejt megkapja a saját DNS másolatát. Nem osztódó sejtek között is lehetséges DNS átvitel ("csupasz" DNS befogásával, bakteriofágok használatával, vagy konjugációval, amikor a baktériumok kopulációs fimbriákkal kapcsolódnak egymáshoz), de ez nem növeli az egyedek számát. A szaporodást a napsugarak és saját létfontosságú tevékenységük termékei akadályozzák.
A baktériumok viselkedése nem különösebben bonyolult. A kémiai receptorok rögzítik a környezet savasságának és a különféle anyagok koncentrációjának változásait: cukrok, aminosavak, oxigén. Sok baktérium reagál a hőmérséklet vagy a fény változására, és néhány baktérium érzékeli a Föld mágneses terét. Kedvezőtlen körülmények között a baktériumot sűrű héj borítja, a citoplazma kiszárad, élettevékenysége szinte megszűnik. Ebben az állapotban a baktériumspórák órákig mély vákuumban maradhatnak, és elviselik a –240 °C és +100 °C közötti hőmérsékletet.
A mikroorganizmusok (mikrobák) 0,1 mm-nél kisebb méretű egysejtűek, amelyek szabad szemmel nem láthatók. Ide tartoznak a baktériumok, mikroalgák, néhány alsóbbrendű fonalas gomba, élesztő és protozoa (1. ábra). A mikrobiológia tanulmányozza őket.
Rizs. 1. Mikrobiológiai objektumok.
ábrán. 2. Az egysejtű protozoák néhány képviselője látható. Néha ennek a tudománynak a tárgyai közé tartoznak a Föld legprimitívebb szervezetei - olyan vírusok, amelyek nem rendelkeznek sejtszerkezettel, és nukleinsavak (genetikai anyag) és fehérje komplexei. Gyakrabban elkülönítik őket egy teljesen külön kutatási területre (Virológia), mivel a mikrobiológia inkább a mikroszkopikus egysejtű szervezetek tanulmányozására irányul.

Rizs. 2. Az egysejtű eukarióták (protozoák) egyes képviselői.
Az algák és gombák vizsgálatával foglalkozó algológia és mikológia tudományok külön tudományágak, amelyek átfedésben vannak a mikrobiológiával a mikroszkopikus élő objektumok vizsgálatában. A bakteriológia a mikrobiológia igazi ága. Ez a tudomány kizárólag a prokarióta mikroorganizmusok vizsgálatával foglalkozik (3. ábra).

Rizs. 3. Prokarióta sejt vázlata.
Az eukariótáktól eltérően, amelyek magukban foglalják az összes többsejtű szervezetet, valamint a protozoonokat, mikroszkopikus algákat és gombákat, a prokariótáknak nincs kialakult magja, amely genetikai anyagot és valódi organellumokat (a sejt állandó speciális struktúráit) tartalmazna.
A prokarióták közé tartoznak a valódi baktériumok és archaeák, amelyeket a modern besorolás szerint doméneknek (szuperkirályságoknak) neveznek Archaea és Eubacteria (4. ábra).

Rizs. 4. A modern biológiai osztályozás területei.
A baktériumok fontos láncszemei a természetben előforduló anyagok körforgásában, lebontják a növényi és állati maradványokat, megtisztítják a szerves anyagokkal szennyezett víztesteket, módosítják a szervetlen vegyületeket. Nélkülük nem létezhetne élet a földön. Ezek a mikroorganizmusok mindenhol elterjedtek, talajban, vízben, levegőben, állati és növényi szervezetekben.
A baktériumok a következő morfológiai jellemzőkben különböznek egymástól:
A gömbölyű baktériumok (coccusok) által alkotott struktúrák között vannak olyan sejtek, amelyek az osztódás után párban állnak, majd egyes képződményekre bomlanak (mikrokokkuszok), vagy folyamatosan együtt maradnak (diplococcusok). A négy sejtből álló négyzetes szerkezetet a tetracoccusok, a láncot a streptococcusok, a 8-64 egységből álló granulátumot a sarcina, a klasztereket a staphylococcusok alkotják.
A pálcika alakú baktériumok változatos alakban jelennek meg a sejt hossza (0,1-15 µm) és vastagsága (0,1-2 µm) nagy eltérése miatt. Ez utóbbi alakja attól is függ, hogy a baktériumok képesek-e spórákat képezni - olyan vastag héjú szerkezeteket, amelyek lehetővé teszik a mikroorganizmusok számára, hogy túléljék a kedvezőtlen körülményeket. Az ezzel a képességgel rendelkező sejteket bacilusoknak nevezzük, azokat pedig, amelyek nem rendelkeznek ilyen tulajdonságokkal, egyszerűen pálcika alakú baktériumoknak.
A pálcika alakú baktériumok speciális módosulatai a fonalas (megnyúlt) formák, láncok és elágazó szerkezetek. Ez utóbbit az aktinomicéták képezik egy bizonyos fejlődési szakaszban. Az „ívelt” rudakat hullámos baktériumoknak nevezik, amelyek között vibriókat izolálnak; két ívű spirilla (15-20 µm); hullámos vonalakra emlékeztető spirocheták. Sejthosszuk 1-3, 15-20 és 20-30 µm. ábrán. Az 5. és 6. ábra a baktériumok főbb morfológiai formáit, valamint a sejtben a spórák elrendezésének típusait mutatja be.

Rizs. 5. A baktériumok alapformái.

Rizs. 6. Baktériumok a sejtben a spórák elhelyezkedésének típusa szerint. 1, 4 – középen; 2, 3, 5 – véghely; 6 – oldalról.
A baktériumok fő sejtszerkezetei: nukleoid (genetikai anyag), fehérjeszintézisre szánt riboszómák, citoplazmatikus membrán (a sejtmembrán része), amely sok képviselőben felülről is védett, kapszula és nyálkahártya (7. ábra).

Rizs. 7. Egy baktériumsejt vázlata.
A baktériumok osztályozása szerint több mint 20 típus létezik. Például rendkívül termofil (magas hőmérsékletet kedvelők) Aquificae, anaerob pálca alakú baktériumok Bacteroidetes. A legdominánsabb törzs azonban, amely változatos képviselőket tartalmaz, az Actinobacteria. Ide tartoznak a bifidobaktériumok, a laktobacillusok és az aktinomicéták. Ez utóbbi egyedisége abban rejlik, hogy egy bizonyos fejlődési szakaszban micéliumot képez.
A köznyelvben ezt micéliumnak hívják. Valójában az aktinomyceták elágazó sejtjei gombahifákra hasonlítanak. Ennek ellenére az aktinomicétákat a baktériumok közé sorolják, mivel prokarióták. Természetesen sejtjeik szerkezetükben kevésbé hasonlítanak a gombákhoz.
Az Actinomycetes (8. ábra) lassan növekvő baktériumok, ezért nem képesek versenyezni a könnyen hozzáférhető szubsztrátumokért. Képesek lebontani olyan anyagokat, amelyeket más mikroorganizmusok nem tudnak szénforrásként felhasználni, különösen a kőolaj-szénhidrogéneket. Ezért az aktinomicétákat intenzíven tanulmányozzák a biotechnológia területén.
Egyes képviselők az olajmezők területére koncentrálnak, és speciális bakteriális szűrőt hoznak létre, amely megakadályozza a szénhidrogének behatolását a légkörbe. Az aktinomyceták gyakorlatilag értékes vegyületek aktív termelői: vitaminok, zsírsavak, antibiotikumok.

Rizs. 8. Reprezentatív aktinomyceta Nocardia.
A mikrobiológia tárgya csak az alsóbbrendű penészgombák (különösen a rhizopus, mucor). Mint minden gomba, nem képes önmagukban anyagokat szintetizálni, és tápközeget igényel. Ennek a királyságnak az alsóbb képviselőinek micéliuma primitív, nincs válaszfalakkal osztva. A mikrobiológiai kutatásban egy speciális rést foglal el az élesztő (9. ábra), amelyet a micélium hiánya jellemez.

Rizs. 9. Élesztőtenyészetek telepformái táptalajon.
Jelenleg sok ismeretet gyűjtöttek össze jótékony tulajdonságaikról. Az élesztőt azonban továbbra is tanulmányozzák gyakorlatilag értékes szerves vegyületek szintézisére való képessége miatt, és aktívan használják modellszervezetként a genetikai kísérletekben. Az ókor óta az élesztőt az erjesztési folyamatokban használták. Az anyagcsere a különböző képviselők között eltérő. Ezért egyes élesztők alkalmasabbak egy adott folyamatra, mint mások.
Például a Saccharomyces beticust, amely jobban ellenáll a magas alkoholkoncentrációnak, erős borok készítésére használják (akár 24%). Míg a S. cerevisiae élesztő kisebb koncentrációjú etanolt képes előállítani. Felhasználási területük szerint az élesztőket takarmányozásra, pékekre, sörfőzdékre, szeszes italokra és borokra osztják.
Betegséget okozó vagy kórokozó mikroorganizmusok mindenhol megtalálhatók. A jól ismert vírusok: influenza, hepatitis, kanyaró, HIV stb. mellett veszélyes mikroorganizmusok a rickettsia, valamint a streptococcusok és a staphylococcusok, amelyek vérmérgezést okoznak. A pálcika alakú baktériumok között számos kórokozó található. Például diftéria, tuberkulózis, tífusz (10. ábra). Az emberre veszélyes mikroorganizmusok számos képviselője megtalálható a protozoonok között, különösen a maláriás plazmódium, a toxoplazma, a leishmania, a lamblia, a trichomonas és a patogén amőbák.

Rizs. 10. Fénykép a lépfenét okozó Bacillus anthracis baktériumról.
Sok aktinomyceta nem veszélyes emberre és állatra. A tuberkulózist és leprát okozó mikobaktériumok között azonban számos patogén képviselő található. Egyes aktinomicéták olyan betegséget indítanak el, mint például az aktinomikózis, amelyet granulomák képződése és néha a testhőmérséklet emelkedése kísér. A penészgombák bizonyos típusai képesek az emberre mérgező anyagokat - mikotoxinokat - termelni. Például az Aspergillus nemzetség néhány képviselője, a Fusarium. A kórokozó gombák a mikózisoknak nevezett betegségek csoportját okozzák. Így a candidiasist vagy egyszerűen fogalmazva a rigót élesztőszerű gombák okozzák (11. ábra). Mindig jelen vannak az emberi szervezetben, de csak akkor aktiválódnak, ha az immunrendszer legyengül.

Rizs. 11. A Candida gomba a rigó kórokozója.
A gombák különféle bőrelváltozásokat okozhatnak, különösen mindenféle zuzmót, kivéve a herpes zoster-t, amelyet vírus okoz. A Malassezia élesztő, az emberi bőr állandó lakója, csökkentheti az immunrendszer aktivitását. Ne rohanjon azonnal kezet mosni. Az egészséges élesztők és opportunista baktériumok fontos funkciót töltenek be, megakadályozva a kórokozók kialakulását.
A vírusok a legprimitívebb élőlények a Földön. Szabad állapotban nem mennek végbe bennük anyagcsere-folyamatok. A vírusok csak akkor kezdenek el szaporodni, amikor belépnek a gazdasejtbe. Minden élő szervezetben a genetikai anyag hordozója a dezoxiribonukleinsav (DNS). Csak a vírusok között vannak olyan genetikai szekvenciával rendelkező képviselők, mint a ribonukleinsav (RNS).
A vírusokat gyakran nem az igazán élő szervezetek közé sorolják.
A vírusok morfológiája igen változatos (12. ábra). Átmérőjük jellemzően 20-300 nm.

Rizs. 12. A vírusrészecskék sokfélesége.
Egyes képviselők 1-1,5 mikron hosszúságot érnek el. A vírus szerkezete abból áll, hogy a genetikai anyagot egy speciális fehérjekerettel (kapszid) veszi körül, amelyet sokféle forma (spirál, ikozaéder, gömb) jellemez. Egyes vírusok tetején a gazdasejt membránjából (szuperkapszid) képződött burok is található. Például (13. ábra) az úgynevezett (AIDS) betegség kórokozójaként ismert. Genetikai anyagként RNS-t tartalmaz, és hatással van az immunrendszer bizonyos típusú sejtjére (segítő T-limfocitákra).

Rizs. 13. A humán immundeficiencia vírus felépítése.
Mikroorganizmusok- ezek kis méretük miatt szabad szemmel láthatatlan élőlények.
A méretkritérium az egyetlen, amely egyesíti őket.
Egyébként a mikroorganizmusok világa még sokszínűbb, mint a makroorganizmusok világa.
A növényekhez és állatokhoz hasonlóan a mikroorganizmusok nevét használják bináris nómenklatúra, azaz általános és konkrét név.
Ha a kutatók nem tudják meghatározni a faji hovatartozást, és csak a nemzetségi hovatartozást határozzák meg, akkor a faj kifejezést használják. Leggyakrabban ez akkor fordul elő, amikor olyan mikroorganizmusokat azonosítanak, amelyek nem hagyományos táplálkozási szükségletekkel vagy életkörülményekkel rendelkeznek. Nemzetség neveáltalában vagy a megfelelő mikroorganizmus morfológiai jellemzői alapján (Staphylococcus, Vibrio, Mycobacterium), vagy a kórokozót felfedező vagy tanulmányozó szerző nevéből származik (Neisseria, Shigella, Escherichia, Rickettsia, Gardnerella).
Faj neve gyakran az e mikroorganizmus által okozott fő betegség nevével (Vibrio cholerae - kolera, Shigella dysenteriae - vérhas, Mycobacterium tuberculosis - tuberculosis) vagy a fő élőhellyel (Escherihia coli - E. coli) társul.
Ezenkívül az orosz nyelvű orvosi irodalomban lehetőség van a baktériumok megfelelő oroszosított elnevezésének használatára (a Staphylococcus epidermidis - epidermális staphylococcus; Staphylococcus aureus - Staphylococcus aureus stb. helyett).
magában foglalja a cianobaktériumok osztályát és az eubaktériumok osztályát, amelyek viszont osztvamegrendelések:
A rendelések csoportokra vannak osztva.
Leghíresebb:
Konjugatív transzfert biztosító F-plazmidok
baktériumok között;
Az R-plazmidok olyan gyógyszerrezisztencia-plazmidok, amelyek biztosítják a különböző betegségek kezelésére használt kemoterápiás szerekkel szembeni rezisztenciát meghatározó gének keringését a baktériumok között.
Prokarióta, túlnyomórészt egysejtű mikroorganizmusok, amelyek hasonló sejtek társulásait (csoportjait) is alkothatják, sejtes, de nem szervezeti hasonlóságokkal jellemezve.
A csoportok további megkülönböztetése családokra, nemzetségekre és fajokra, amelyek a fő taxonómiai kategória, a biokémiai tulajdonságok vizsgálata alapján történik. Ez az elv képezi a baktériumok speciális kézikönyvekben megadott osztályozásának alapját - baktériumok meghatározói.
Kilátás Egy genotípusú egyedek evolúciósan kialakult halmaza, amely standard körülmények között hasonló morfológiai, fiziológiai és biokémiai jellemzőkkel nyilvánul meg.
A kórokozó baktériumok esetében a „faj” definíciója kiegészül azzal a képességgel, hogy bizonyos nozológiai betegségeket okozhatnak.
A mikrobiológiában széles körben használják a speciális kifejezéseket - tenyészet, törzs, klón.
Kultúra a szemmel látható baktériumok gyűjteménye tápközegen.
A tenyészetek lehetnek tiszták (egy fajba tartozó baktériumok halmaza) vagy kevertek (2 vagy több fajból álló baktériumok halmaza).
Szűrd le ugyanazon fajba tartozó baktériumok gyűjteménye, amelyeket különböző forrásokból vagy ugyanabból a forrásból különböző időpontokban izoláltak.
A törzsek bizonyos jellemzőikben különbözhetnek, amelyek nem haladják meg a faj jellemzőit. Klón olyan baktériumok gyűjteménye, amelyek egyetlen sejt utódai.
Az emberek új módszereket próbálnak találni, hogy megvédjék magukat káros befolyásaiktól. De vannak jótékony mikroorganizmusok is: elősegítik a tejszín érését, a növények nitrát képződését, lebontják az elhalt szöveteket stb. A mikroorganizmusok vízben, talajban, levegőben, az élő szervezetek testén és azok belsejében élnek.
A baktériumok alakjai
A baktériumoknak négy fő formája van, nevezetesen:



A baktérium összetett szerkezettel rendelkezik:
Javasoljuk, hogy vegye figyelembe a baktériumok fő típusait:



A tápanyagok a sejt teljes felületén keresztül jutnak be. A mikroorganizmusok széles körben elterjedtek a különféle táplálkozási módok megléte miatt. Életükhöz sokféle elemre van szükségük: szénre, foszforra, nitrogénre stb. A tápanyagellátást membrán szabályozza.
A táplálkozás típusát a szén és a nitrogén felszívódásának módja és az energiaforrás típusa határozza meg. Egyesek ezeket az elemeket a levegőből nyerhetik, és napenergiát használhatnak, míg mások szerves eredetű anyagokat igényelnek. Mindannyiuknak szükségük van vitaminokra és aminosavakra, amelyek katalizátorként működhetnek a szervezetükben fellépő reakciókban. Az anyagok eltávolítása a sejtből a diffúziós folyamaton keresztül történik.
Sokféle mikroorganizmusban az oxigén fontos szerepet játszik az anyagcserében és a légzésben. A légzés hatására energia szabadul fel, amit szerves vegyületek előállítására használnak fel. De vannak olyan baktériumok, amelyek számára az oxigén halálos.
A szaporodás a sejt két részre osztásával történik. Miután elér egy bizonyos méretet, megkezdődik az elválasztási folyamat. A sejt megnyúlik, és keresztirányú septum képződik benne. A keletkező részek szétszóródnak, de egyes fajok kapcsolatban maradnak, és klasztereket alkotnak. Az újonnan kialakult részek mindegyike önálló szervezetként táplálkozik és nő. Kedvező környezetbe helyezve a szaporodási folyamat nagy sebességgel megy végbe.

A mikroorganizmusok képesek az összetett anyagokat egyszerű anyagokra bontani, amelyeket aztán újra felhasználhatnak a növények. Ezért a baktériumok nélkülözhetetlenek az anyagok körforgásában, nélkülük sok fontos folyamat lehetetlen lenne a Földön.
Tudod?

Következtetés: Ne felejtsen el kezet mosni minden alkalommal, amikor hazatér, miután kiment a szabadba. Ha vécére megy, mosson kezet is szappannal. Egyszerű szabály, de nagyon fontos! Tartsa tisztán, és a baktériumok nem fognak zavarni!
Az anyag megerősítése érdekében meghívjuk Önt izgalmas feladataink teljesítésére. Sok szerencsét!
1. számú feladat
Nézze meg figyelmesen a képet, és mondja meg, melyik sejt bakteriális? Próbálja meg elnevezni a fennmaradó cellákat anélkül, hogy megnézné a nyomokat:

A baktériumok osztályozása. A baktériumok nemzetközi kódexének határozata a következő taxonómiai kategóriákat javasolta: osztály, osztály, rend, család, nemzetség, faj. A fajnév bináris nómenklatúrának felel meg, azaz két szóból áll. Például a szifilisz kórokozóját úgy írják le Treponema pallidum. Az első szó na-
a nemzetség nevét és nagybetűvel írják, a második szó a fajt jelöli és kisbetűvel írják. Ha egy fajt ismét megemlítenek, a generikus név a kezdőbetűre rövidül, például: T.pallidum.
A baktériumok prokarióták, azaz. prenukleáris szervezetek, mivel primitív magjuk van héj, nukleolusz vagy hisztonok nélkül. és a citoplazmából hiányoznak a jól szervezett organellumok (mitokondriumok, Golgi-készülék, lizoszómák stb.)
A régi Bergey-féle szisztematikus bakteriológiai kézikönyvben a baktériumokat a bakteriális sejtfal jellemzői szerint 4 részre osztották: Gracilicutes - vékony sejtfalú, gram-negatív eubaktériumok; Cégek - vastag sejtfalú eubaktériumok, gram-pozitívak; Tenericutes - sejtfal nélküli eubaktériumok; Mendosicutes - hibás sejtfalú archaebaktériumok.
Minden osztályt szekciókra vagy csoportokra osztottak a Gram-festés, a sejtforma, az oxigénigény, a motilitás, az anyagcsere és a táplálkozási jellemzők alapján.
A kézikönyv 2. kiadása (2001) szerintBergey, a baktériumok 2 doménre oszlanak:"Baktériumok" és "Archaea" (2.1. táblázat).
Asztal. Domain jellemzőiBaktériumokÉsArchaea
|
Tartomány"baktériumok"(eubaktériumok) |
Tartomány"Archaea" (archebacteriumok) |
|
A „baktériumok” tartományban megkülönböztethetünk a következő baktériumok: 1) vékony sejtfalú baktériumok, gram-negatívak*; 2) vastag sejtfalú baktériumok, gram-pozitívak**; 3) béta-sejtfalbaktériumok (Mollicutes osztály – mikoplazmák) |
Az archaebaktériumok sejtfalukban nem tartalmaznak peptidoglikánt. Speciális riboszómáik és riboszomális RNS-ük (rRNS) van. Az „archebacteria” kifejezés 1977-ben jelent meg. Ez az élet egyik ősi formája, amint azt az „arche” előtag is jelzi. Nincs köztük fertőző ágens |
*Vékony falú gram-negatív eubaktériumok között megkülönböztetni:
gömb alakú formák vagy coccusok (gonococcusok, meningococcusok, veillonella);
csavart formák - spirochetes és spirilla;
rúd alakú formák, beleértve a rickettsiát is.
** Vastag falú gram-pozitív eubaktériumokhoz tartalmazza:
gömb alakú formák vagy coccusok (staphylococcusok, streptococcusok, pneumococcusok);
rúd alakú formák, valamint aktinomyceták (elágazó, fonalas baktériumok), corynebacteriumok (klub alakú baktériumok), mikobaktériumok és bifidobaktériumok (2.1. ábra).
A legtöbb gram-negatív baktérium a Proteobaktériumok törzsébe tartozik. a riboszómális RNS hasonlósága alapján „Proteobaktériumok” – a görög Proteus istenről nevezték el. különböző formákat öltve). Közös fotoszintézisből jelentek meg tic őse.
A Gram-pozitív baktériumok a vizsgált riboszómális RNS-szekvenciák szerint külön filogenetikai csoportot alkotnak, két nagy alosztályból - magas és alacsony arányban. G+ C (genetikai hasonlóság). A Proteobaktériumokhoz hasonlóan ez a csoport metabolikusan változatos.
Domainhez "Baktériumok» 22 típust tartalmaz, ebbőlA következők nagyon fontosak az orvostudományban:
típusProteobaktériumok
Osztály Alfaproteobaktériumok. Szülés: Rickettsia, Orientia, Ehrlichia, Bartonella, Brucella
Osztály Betaproteobaktériumok. Szülés: Burkholderia, Alcaligenes, Bordetella, Neisseria, Kingella, Spirillum
Osztály Gammaproteobaktériumok. Szülés: Francisella, Legionella, Coxiella, Pseudomonas, Moraxella, Acinetobacter, Vibrio, Enterobacter, Callimatobacterium, Citrobacter, Edwardsiella, Erwinia, Escherichia, Hafnia, Klebsiella, Morganella, Proteus, Providencia, Salmonella, Serratia, Pasteureers, Shigella
Osztály Deltaproteobaktériumok. Nemzetség: Bilophila
Osztály Epszilonproteobaktériumok. Szülés: Campylobacter, Helicobacter, Wolinella
típusFirmikusok (fő-útgrampolo lakos)
Osztály Clostridia. Szülés: Clostridium, Sarcina, Peptostreptococcus, Eubacterium, Peptococcus, Veillonella (Gram-negatív)
Osztály Mollicutes. Nemzetek: Mycoplasma, Ureaplasma
Osztály Bacillusok. Szülés: Bacillus, Sporosarcina, Listeria, Staphylococcus, Gemella, Lactobacillus, Pediococcus, Aerococcus, Leuconostoc, Streptococcus, Lactococcus
típusActinobacteriumok
Osztály Actinobacteriumok. Szülés: Actinomyces, Arcanodacterium, Mobiluncus, Micrococcus, Rothia, Stomatococcus, Corynebacterium, Mycobacterium, Nocardia, Propionibacterium, Bifidobacterium, Gardnerella
típusClamydiae
Osztály Clamydiae. Szülés: Clamydia, Clamydophila
típusSpirocheták
Osztály Spirocheták. Szülés: Spirochaeta, Borrelia, Treponema, Leptospira
Phylum Bacteroidetes
Osztály Bakterioidák. Szülés: Bacteroides, Porphyromonas, Prevotella
Osztály Flavobaktériumok. Szülés: Flavobacterium
A baktériumok sejtfal szerkezeti sajátosságai szerinti osztódása a Gram-módszerrel színezésük lehetséges változatosságával függ össze. E módszer szerint, amelyet H. Gram dán tudós 1884-ben javasolt, a festési eredményektől függően a baktériumokat Gram-pozitív, kék-lilára festett és Gram-negatív, vörösre festett baktériumokra osztják. Kiderült azonban, hogy az úgynevezett Gram-pozitív típusú (vastagabb, mint a Gram-negatív baktériumoké) sejtfalú baktériumok, például a Mobiluncus nemzetségbe tartozó baktériumok és néhány spóraképző baktérium a szokásos gram helyett. -pozitív szín, Gram-negatív színű. Ezért a baktériumok taxonómiája szempontjából a sejtfalak szerkezeti jellemzői és kémiai összetétele nagyobb jelentőséggel bír, mint a Gram-festés.
2.2.1. A baktériumok alakjai
A baktériumoknak számos fő formája létezik (lásd a 2.1. ábrát) - kokcoid, rúd alakú, tekercses és elágazó, fonalas formái a baktériumoknak.
Gömb alakú formák vagy coccusok,- 0,5-1,0 mikron méretű gömbbaktériumok*, amelyek egymáshoz viszonyított helyzetük szerint mikrococcusokra, diplococcusokra, streptococcusokra, tetracoccusokra, sarcinákra oszlanak És staphylococcusok.
Mikrococcusok(görögből mikros - kicsi) - külön elhelyezkedő sejtek.
Diplococcus(görögből diploos - kettős), vagy páros coccusok, párban helyezkednek el (pneumococcus, gonococcus, meningococcus), mivel a sejtek osztódás után nem válnak el. Pneumococcus (a tüdőgyulladás kórokozója) ellentétes oldalán lándzsa alakú, ill gonococcus(a gonorrhoea kórokozója) és a meningococcus (a járványos agyhártyagyulladás kórokozója) kávébab alakúak, homorú felületük egymás felé néz.
Streptococcusok(görögből streptos - lánc) - kerek vagy hosszúkás sejtek, amelyek láncot alkotnak az azonos síkban történő sejtosztódás és a köztük lévő kapcsolat megőrzése miatt az osztódás helyén.
Sarcins(a lat. sarcina - csomó, bála) 8 vagy több coccusból álló csomagok formájában vannak elrendezve, mivel a sejtosztódás során három egymásra merőleges síkban jönnek létre.
Staphylococcus(görögből stafil - szőlőfürt) - cocci, szőlőfürt formájában rendeződnek el a különböző síkokba való osztás eredményeként.
Rúd alakú baktériumok különböznek méretükben, a sejtvégek alakjában és a sejtek egymáshoz viszonyított helyzetében. A sejtek hossza 1,0-10 µm, vastagsága 0,5-2,0 µm. A rudak lehetnek szabályosak (E. coli stb.) és szabálytalanok (korinebaktériumok). És egyéb) formák, beleértve az elágazó formákat is, például az aktinomicétákban. A legkisebb rúd alakú baktériumok közé tartozik a rickettsia.
A rudak végei levághatók (anthrax bacillus), lekerekítettek (Escherichia coli), hegyesek (fuzobaktériumok) vagy megvastagodás formájában. Ez utóbbi esetben a rúd úgy néz ki, mint egy ütő (Corynebacterium diphtheria).
Az enyhén ívelt rudakat vibrióknak (Vibrio cholerae) nevezik. A legtöbb pálcika alakú baktérium véletlenszerűen helyezkedik el, mivel a sejtek osztódás után eltávolodnak egymástól. Ha a sejtosztódás után a sejtek kapcsolatban maradnak,
Ha a sejtfal közös töredéke van, és nem térnek el egymástól, akkor egymással szöget zárnak be (Corynebacterium diphtheria), vagy láncot alkotnak (antrax bacillus).
Csavart formák- például spirál alakú baktériumok spirilla, dugóhúzó alakú csavart cellák megjelenésével. A kórokozó spirilla magában foglalja a sodoku (patkányharapásos betegség) kórokozóját. A tekervényesek közé tartozik a Campilobacter és a Helicobacter is, amelyek rendelkeznek kanyarodik mint a repülő sirály szárnya; a baktériumok, például a spirocheták is közel állnak hozzájuk. Spirochetes- vékony, hosszú, hullámos
spirál alakú) baktériumok, amelyek a sejtek hajlítási változásai miatt mobilitásban különböznek a spirillától. A spirocheták egy külső membránból állnak
sejtfal) protoplazma hengert körülvevő citoplazma membránnal és axiális filamentummal (axistil). Az axiális filamentum a sejtfal külső membránja alatt helyezkedik el (a periplazmában), és mintegy a spirocheta protoplazmatikus hengere körül csavarodik, így spirális formát ad (a spirocheta elsődleges fürtjei). Az axiális filamentum periplazmatikus fibrillákból áll - a bakteriális flagella analógjaiból, és egy összehúzódó flagellin fehérje. A fibrillák a sejt végeihez kapcsolódnak (2.2. ábra), és egymás felé irányulnak. A fibrillák másik vége szabad. A fibrillumok száma és elrendezése fajonként változó. A rostok részt vesznek a spirocheták mozgásában, így a sejteknek rotációs, hajlító és transzlációs mozgást adnak. Ebben az esetben a spirocheták hurkokat, fürtöket és hajlatokat képeznek, amelyeket másodlagos fürtöknek neveznek. Spirochetes
nem fogadja jól a festékeket. Általában Romanovsky-Giemsa szerint festettek vagy ezüstözöttek. Az élő spirocétákat fáziskontrasztos vagy sötétmezős mikroszkóppal vizsgáljuk.
A spirochetákat 3 nemzetség képviseli, amelyek patogén az emberre: Treponema, Borrelia, Leptospira.
Treponema(Treponema nemzetség) vékony, dugóhúzóval csavart szálak, 8-12 egységes kis fürtökkel. A treponema protoplasztja körül 3-4 rost (flagella) található. A citoplazma citoplazmatikus szálakat tartalmaz. A kórokozók képviselői T.pallidum - a szifilisz kórokozója, T.pertenue - a trópusi betegség kórokozója. Vannak szaprofiták is - az emberi szájüreg lakói és a tározók iszapja.
Borrelia(nemzetség Borrelia), a treponemákkal ellentétben hosszabbak, 3-8 nagy fürtjük és 7-20 rostjuk van. Ezek közé tartozik a visszaeső láz kórokozója (BAN BEN.recurrentis) és a Lyme-kór kórokozói (BAN BEN.burgdorferi satöbbi.).
Leptospira(nemzetség Leptospira) Sekély és gyakori fürtök vannak - csavart kötél formájában. Ezeknek a spirochetáknak a végei ívesek, mint a horgok, a végén vastagodással. Másodlagos fürtöket képezve betűk megjelenését veszik fel S vagy együtt; 2 axiális szála van (flagella). A kórokozó képviselője L. ban ben terrogánok vízzel vagy étellel lenyelve leptospirózist okoz, ami vérzések és sárgaság kialakulásához vezet.
a citoplazmában, néhány pedig a fertőzött sejtek magjában. Ízeltlábúakban (tetvekben, bolhákban, kullancsokban) élnek, amelyek gazdáik vagy hordozóik. A Rickettsia nevét H. T. Ricketts amerikai tudósról kapta, aki először írta le az egyik kórokozót (Sziklás hegyi foltos láz). A rickettsia alakja és mérete a növekedési körülményektől függően változhat (szabálytalan, fonalas sejtek). A rickettsia szerkezete nem különbözik a gram-negatív baktériumokétól.
A riketsiák anyagcseréje független a gazdasejttől, azonban lehetséges, hogy a szaporodáshoz nagy energiájú vegyületeket kapnak a gazdasejttől. A kenetekben és a szövetekben Romanovsky-Giemsa, Macchiavello-Zdrodovsky szerint festettek (a rickettsiák vörösek, a fertőzött sejtek kékek).
Emberben a rickettsiák járványos tífuszt okoznak. (Rickettsia prowazekii), kullancs által terjesztett rickettsiosis (R. sibirica), Rocky Mountain foltos láz (R. rickettsii) és egyéb rickettsiosisok.
Az elemi testek endocitózissal jutnak be a hámsejtekbe, intracelluláris vakuólum képződésével. A sejtek belsejében megnagyobbodnak és osztódó retikuláris testekké alakulnak át, és vakuolákban (zárványokban) fürtöket képeznek. Az elemi testek retikuláris testekből jönnek létre, amelyek exocitózissal vagy sejtlízissel hagyják el a sejteket. Akik elmentek
Az elemi testsejtek új ciklusba lépnek, megfertőzve más sejteket (16.11.1. ábra). Emberben a chlamydia károsítja a szemet (trachoma, kötőhártya-gyulladás), az urogenitális traktusban, a tüdőben stb.
Actinomycetes- elágazó, fonalas vagy rúd alakú Gram-pozitív baktériumok. A neve (görögből. actis - Ray, mykes - gomba) kaptak az érintett szövetekben drusen képződése miatt - szorosan összefonódó szálak szemcséi, amelyek sugarak formájában terjednek ki a központból és lombik alakú megvastagodásokkal végződnek. Az aktinomyceták a gombákhoz hasonlóan micéliumot - fonalszerűen összefonódó sejteket (hifákat) alkotnak. Szubsztrát micéliumot képeznek, amely a sejt tápközegbe való benövése eredményeként jön létre, és légmicéliumot, amely a táptalaj felületén nő. Az aktinomyceták a micélium feldarabolásával a pálcika- és cocci alakú baktériumokhoz hasonló sejtekre osztódhatnak. Az aktinomyceták légi hifáin spórák képződnek, amelyek a szaporodást szolgálják. Az Actinomycete spórái általában nem hőállóak.
Az aktinomicétákkal közös filogenetikai ágat az úgynevezett nocardi-szerű (nocardioform) aktinomicéták alkotják, amelyek a rúd alakú, szabálytalan alakú baktériumok kollektív csoportja. Egyéni képviselőik elágazó formákat alkotnak. Ide tartoznak a nemzetséghez tartozó baktériumok Corynebacterium, Mycobacterium, Nocardianjxp. A Nocardi-szerű aktinomicétákat az jellemzi, hogy a sejtfalban arabinóz, galaktóz, valamint mikolsav és nagy mennyiségű zsírsav található a sejtfalban. A mikolsavak és a sejtfallipidek meghatározzák a baktériumok savállóságát, különösen a Mycobacterium tuberculosis és a lepra (Ziehl-Neelsen szerint festve vörös, a nem saválló baktériumok és szövetelemek, a köpet pedig kék színűek).
A kórokozó aktinomicéták aktinomikózist, nocardiát - nocardiózist, mycobacteriumot - tuberkulózist és leprát, korinebaktériumot - diftériát okoznak. A talajban elterjedtek az aktinomyceták szaprofita formái és a nocardia-szerű aktinomyceták, sok közülük antibiotikum termelő.
Sejtfal- erős, rugalmas szerkezet, amely a baktériumnak bizonyos formát ad, és az alatta lévő citoplazmatikus membránnal együtt „fékezi” a baktériumsejtben a magas ozmotikus nyomást. Részt vesz a sejtosztódás folyamatában és a metabolitok szállításában, receptorai vannak bakteriofágokhoz, bakteriocinokhoz és különféle anyagokhoz. A legvastagabb sejtfal a gram-pozitív baktériumokban található (2.4. és 2.5. ábra). Tehát, ha a Gram-negatív baktériumok sejtfalának vastagsága körülbelül 15-20 nm, akkor a Gram-pozitív baktériumokban elérheti az 50 nm-t vagy még többet is.
Mikoplazmák- kisméretű baktériumok (0,15-1,0 µm), amelyeket csak citoplazmatikus membrán vesz körül. Az osztályhoz tartoznak Mollicutes, szterolokat tartalmaznak. A sejtfal hiánya miatt a mikoplazmák ozmotikusan érzékenyek. Különböző formájúak: coccoid, fonalas, lombik alakúak. Ezek a formák a tiszta mikoplazma tenyészetek fáziskontraszt mikroszkópos vizsgálatakor láthatók. Sűrű táptalajon a mikoplazmák tükörtojásra emlékeztető kolóniákat alkotnak: egy központi, átlátszatlan részt a táptalajba merülve, és egy áttetsző kerületet kör alakban.
A mikoplazmák atipikus tüdőgyulladást okoznak emberben (Mycoplasma pneumoniae) és az urogenitális traktus elváltozásai (M.homi- nis satöbbi.). A mikoplazmák nemcsak állatokban, hanem növényekben is betegségeket okoznak. A nem patogén képviselők is meglehetősen elterjedtek.
2.2.2. A bakteriális sejt szerkezete
A baktériumok szerkezetét jól tanulmányozták egész sejtek és vékony metszeteik elektronmikroszkópos vizsgálatával, valamint más módszerekkel. A baktériumsejtet sejtfalból és citoplazmatikus membránból álló membrán veszi körül. A héj alatt protoplazma található, amely zárványokkal ellátott citoplazmából és egy nukleoidnak nevezett magból áll. Vannak további struktúrák: kapszula, mikrokapszula, nyálka, flagella, pili (2.3. ábra). Egyes baktériumok kedvezőtlen körülmények között képesek spórákat képezni.
A gram-pozitív baktériumok sejtfalában kis mennyiségben poliszacharidokat, lipideket és fehérjéket tartalmaz. E baktériumok sejtfalának fő összetevője a többrétegű peptidoglikán (mu-rein, mukopeptid), amely a sejtfal tömegének 40-90%-át teszi ki. Teichoin savak (a görögből. teichos - fal), melynek molekulái 8-50 glicerin- és ribitolmaradékból álló láncok, amelyeket foszfáthidak kötnek össze. A baktériumok alakját és erejét a többrétegű peptidoglikán merev rostos szerkezete adja, amely peptidekkel térhálósodik.
A peptidoglikánt párhuzamos molekulák képviselik glikán. ismétlődő N-acetil-glükózamin- és N-acetil-muraminsav-maradékokból áll, amelyeket glikozidos kötéssel kapcsolnak össze. Ezeket a kötéseket a lizozim, amely egy acetil-muramidáz bontja meg. A glükánmolekulák N-acetil-muraminsavon keresztül kapcsolódnak össze négy aminosavból álló peptid keresztkötéssel ( tetrapeptid). Innen származik a polimer neve - peptidoglikán.
A Gram-negatív baktériumokban a peptidoglikán peptidkötésének alapja a váltakozó L- és D-aminosavakból álló tetrapeptidek, például: L-alanin - D-glutaminsav - mezo-diaminopimelinsav - D-alanin. U E.coli (gram-negatív baktérium) peptidláncok az egyik lánc D-alaninján és a mezo-diaminopimeli-n keresztül kapcsolódnak egymáshoz.
új sav – másik. A Gram-negatív baktériumok peptidoglikánja peptid részének összetétele és szerkezete stabil, ellentétben a Gram-pozitív baktériumok peptidoglikánjával, amelynek aminosavai összetételükben és sorrendjükben eltérőek lehetnek. A Gram-pozitív baktériumok peptidoglikán-tetrapeptidjei 5 aminosavból álló polipeptidláncokkal kapcsolódnak egymáshoz
glicin (pentaglicin). A mezo-diamino-pimelinsav helyett gyakran lizint tartalmaznak. A glükán elemek (acetil-glükózamin és acetil-muraminsav) és a tetrapeptid aminosavak (mezo-diaminopimelinsav és D-glutaminsav, D-alanin) a baktériumok megkülönböztető jellemzői, mivel állatokban és emberekben hiányoznak.
A Gram-pozitív baktériumok azon képessége, hogy megtartsák az enciánibolyát jóddal kombinálva, ha Gram-festéssel (a baktériumok kék-lila színe) festik, a többrétegű peptidoglikánnak a festékkel kölcsönhatásba lépő tulajdonságával függ össze. Ezenkívül a bakteriális kenet utólagos alkoholos kezelése a peptidoglikán pórusainak szűkülését okozza, és ezáltal megtartja a festéket a sejtfalban. A Gram-negatív baktériumok alkohollal való érintkezés után elveszítik a festéket, ami a kisebb mennyiségű peptidoglikánnak köszönhető (a sejtfal tömegének 5-10%-a); alkohollal elszíneződnek, és fukszinnal vagy szafraninnal kezelve vörös színt kapnak.
BAN BEN Gram-negatív baktériumok sejtfalának összetétele bejut a külső membránba, és lipoproteinen keresztül kapcsolódik az alatta lévő peptidoglikán réteghez (2.4. és 2.6. ábra). A baktériumok ultravékony metszeteinek elektronmikroszkópos vizsgálatakor a külső membrán egy hullámos háromrétegű szerkezetnek tűnik, hasonlóan a belső membránhoz, amelyet citoplazmatikusnak neveznek. Ezeknek a membránoknak a fő összetevője egy bimolekuláris (kettős) lipidréteg.
A külső membrán egy mozaik szerkezet, amelyet lipopoliszacharidok, foszfolipidek és fehérjék képviselnek. Belső rétegét foszfolipidek képviselik, a külső réteget pedig tartalmaz lipopoliszacharid(LPS). Így a külső membrán aszimmetrikus. Az LPS külső membrán három részből áll:
lipid A - konzervatív szerkezet, majdnem ugyanaz a gram-negatív baktériumokban;
mag, vagy mag, kéregrész (lat. mag - mag), viszonylag konzervált oligoszacharid szerkezet;
egy nagyon variábilis O-specifikus poliszacharid lánc, amely azonos oligoszacharid szekvenciák ismétlődéséből jön létre.
Az LPS-t a lipid A „rögzíti” a külső membránban, ami az LPS toxicitását okozza, ezért endotoxinnal azonosítják. A baktériumok antibiotikumok általi elpusztítása nagy mennyiségű endotoxin felszabadulásához vezet, ami endotoxikus sokkot okozhat a betegben. Az LPS magja vagy magrésze a lipid A-ból nyúlik ki. Az LPS mag legállandóbb része a keto-dezoxioktonsav (3-dezoxi-O-man-no-2-oktulozonsav). Az LPS-molekula magjából kinyúló O-specifikus lánc meghatározza egy adott baktériumtörzs szerocsoportját, szerovariáját (az immunszérum által kimutatott baktériumtípus). Így az LPS fogalma összefügg az O-antigén fogalmával, amellyel a baktériumok megkülönböztethetők. A genetikai változások hibákhoz, a bakteriális LPS „lerövidüléséhez” és az ebből eredő R-formák „durva” kolóniáihoz vezethetnek.
A külső membrán mátrixfehérjéi áthatolnak rajta oly módon, hogy a porinoknak nevezett fehérjemolekulák határolják a hidrofil pórusokat, amelyeken keresztül víz és kis, legfeljebb 700 Da relatív tömegű hidrofil molekulák haladnak át.
A külső és a citoplazmatikus membrán között egy periplazmatikus tér vagy periplazma található, amely enzimeket (proteázokat, lipázokat, foszfatázokat,
nukleázok, béta-laktamázok), valamint a transzportrendszerek összetevői.
Ha a bakteriális sejtfal szintézise lizozim, penicillin, a test védőfaktorai és más vegyületek hatására megszakad, módosított (gyakran gömb alakú) sejtek képződnek: protoplasztok - sejtfaltól teljesen mentes baktériumok; A szferoplasztok részben megőrzött sejtfalú baktériumok. A sejtfal-inhibitor eltávolítása után az ilyen megváltozott baktériumok megfordulhatnak, azaz teljes sejtfalat kaphatnak, és visszaállíthatják eredeti alakjukat.
Azokat a szfero- vagy protoplaszt típusú baktériumokat, amelyek elvesztették a peptidoglikán szintézisének képességét antibiotikumok vagy más tényezők hatására, és képesek szaporodni, L-formáknak nevezik (a D. Lister Institute nevéből, ahol voltak először tanulmányozta). Az L-formák mutációk következtében is kialakulhatnak. Ozmotikusan érzékeny, gömb alakú, lombik alakú, különböző méretű sejtek, beleértve a baktériumszűrőkön átmenőket is. Egyes L-formák (instabilok), ha a baktériumok változását okozó tényezőt eltávolítják, megfordulhatnak, és „visszatérhetnek” az eredeti baktériumsejthez. L-formákat számos fertőző betegség kórokozója képes előállítani.
Citoplazma membrán ana ultravékony metszetek elektronmikroszkópos vizsgálatánál háromrétegű membrán (2 sötét réteg, egyenként 2,5 nm vastag, egy világos köztes réteg választja el). Szerkezetében (lásd a 2.5. és 2.6. ábrát) hasonló az állati sejtek plazmalemmájához, és kettős lipidrétegből, főként foszfolipidekből áll, beágyazott felülettel és integrált fehérjékkel, amelyek úgy tűnik, hogy áthatolnak a membrán szerkezetén. Ezek egy része az anyagok szállításában részt vevő permeáz.
A citoplazmatikus membrán egy dinamikus szerkezet mobil komponensekkel, ezért úgy gondolják, mint egy mozgó folyadékszerkezetet. Körülveszi a bakteriális citoplazma külső részét, és részt vesz az ozmotikus nyomás szabályozásában.
niya, anyagszállítás és a sejt energia-anyagcseréje (az elektrontranszport lánc enzimei, adenozin-trifoszfatáz stb. miatt).
A túlzott növekedéssel (a sejtfal növekedéséhez képest) a citoplazmatikus membrán invaginátumokat képez - invaginációk összetett csavart membránszerkezetek, úgynevezett mezoszómák formájában. A kevésbé bonyolultan csavart struktúrákat intracitoplazmatikus membránoknak nevezzük. A mezoszómák és az intracitoplazmatikus membránok szerepe nem teljesen ismert. Még azt is feltételezik, hogy olyan műtermékről van szó, amely az elektronmikroszkópos minta előkészítése (rögzítése) után keletkezik. Mindazonáltal úgy vélik, hogy a citoplazma membrán származékai részt vesznek a sejtosztódásban, energiát adva a sejtfal szintéziséhez, részt vesznek az anyagok kiválasztásában és a spóraképzésben, vagyis a nagy energiafelhasználású folyamatokban.
A citoplazma a baktériumsejt fő térfogatát foglalja el, és oldható fehérjékből, ribonukleinsavakból, zárványokból és számos kis szemcsékből - riboszómákból áll, amelyek a fehérjék szintéziséért (transzlációjáért) felelősek.
A bakteriális riboszómák mérete körülbelül 20 nm, ülepedési együtthatója pedig 70S, ellentétben az eukarióta sejtekre jellemző SOS riboszómákkal. Ezért egyes antibiotikumok a bakteriális riboszómákhoz való kötődés révén gátolják a bakteriális fehérjeszintézist anélkül, hogy befolyásolnák az eukarióta sejtekben a fehérjeszintézist. A bakteriális riboszómák két alegységre - 50S és 30S - disszociálhatnak. A riboszómális RNS-ek (rRNS-ek) a baktériumok konzervált elemei (az evolúció „molekuláris órája”). A 16S rRNS a kis riboszomális alegység, a 23S rRNS pedig a nagy riboszomális alegység része. A 16S rRNS vizsgálata a génrendszertani alapja, lehetővé téve az élőlények rokonsági fokának felmérését.
A citoplazma különféle zárványokat tartalmaz glikogén granulátum, poliszacharidok, béta-hidroxi-vajsav és polifoszfátok (volutin) formájában. Felhalmozódnak, ha tápanyagfelesleg van a környezetben és
Tartalékanyagként működnek a táplálkozásban és az energiaszükségletben.
A Volutin affinitást mutat az alapfestékekhez, és könnyen kimutatható speciális festési módszerekkel (például Neisser) metakromatikus granulátum formájában. A toluidin kékkel vagy metilénkékkel a volutin vörös-ibolyára festődik, a baktérium citoplazmája pedig kékre festődik. A diftéria bacillusban a volutin szemcsék jellegzetes elrendeződése intenzíven festődő sejtpólusok formájában mutatkozik meg. A volutin metakromatikus festődése a polimerizált szervetlen polifoszfát magas tartalmához kapcsolódik. Elektronmikroszkóppal úgy néznek ki, mint a 0,1-1,0 mikron méretű elektronsűrű granulátumok.
Nukleoid- egyenértékű a baktériumok sejtmagjával. A baktériumok központi zónájában található, kettős szálú DNS formájában, gyűrűbe zárva és szorosan, mint egy labda. A baktériumok magjában, az eukariótáktól eltérően, nincs nukleáris burok, sejtmag és bázikus fehérjék (hisztonok). A baktériumsejt jellemzően egy kromoszómát tartalmaz, amelyet egy gyűrűbe zárt DNS-molekula képvisel, és ha az osztódás megszakad, 4 vagy több kromoszóma konvergálhat benne. A nukleoidot fénymikroszkóppal detektálják festés után DNS-specifikus módszerekkel: Feulgen vagy Romanovsky-Giemsa. A baktériumok ultravékony metszeteinek elektrondiffrakciós mintázataiban a nukleoid világos zónákként jelenik meg, fibrilláris, fonalszerű DNK-struktúrákkal, amelyek bizonyos területekhez kötődnek.
citoplazma membrán vagy mezozo-
az enyém, részt vesz a kromoszóma replikációjában (lásd 2.5. és 2.6. ábra).
A nukleoidon kívül, amelyet egy képvisel
kromoszóma, baktériumsejtben vannak
az öröklődés kromoszómán kívüli tényezői -
plazmidok (lásd az 5.1.2. pontot), amelyek reprezentálják
a DNS kovalensen zárt gyűrűi.
Kapszula, mikrokapszula, nyálka . Kapszula-
0,2 mikronnál vastagabb nyálkahártya szerkezet, amely szorosan kapcsolódik a baktériumok sejtfalához, és világosan meghatározott külső határokkal rendelkezik. A kapszula kóros anyagból származó ujjlenyomat-kenetekben látható. A tiszta baktériumkultúrákban kapszula képződik
ritkábban. Speciális Burri-Gins szerinti kenetfestési módszerekkel mutatják ki, ami negatív kontrasztot hoz létre a kapszula anyagai között: a tinta sötét hátteret hoz létre a kapszula körül.
A kapszula poliszacharidokból (exopoliszacharidokból), néha polipeptidekből áll; például az anthrax bacillusban D-glutaminsav polimerjeiből áll. A kapszula hidrofil és nagy mennyiségű vizet tartalmaz. Megakadályozza a baktériumok fagocitózisát. Kapszula antigén-na: A kapszula elleni antitestek okozzák növekedés (duzzadási reakcióés én kapszula ly).
Sok baktérium mikrokapszulát alkot – 0,2 mikronnál kisebb vastagságú nyálkahártya-képződményt, amely csak elektronmikroszkóppal detektálható. A nyálkát meg kell különböztetni a kapszulától - nyálkahártya exopoliszacharidoktól, amelyeknek nincs egyértelmű külső határa. A nyálka vízben oldódik.
A mukoid exopoliszacharidok a Pseudomonas aeruginosa nyálkahártya törzseire jellemzőek, gyakran megtalálhatók a cisztás fibrózisban szenvedő betegek köpetében. A bakteriális exopoliszacharidok részt vesznek az adhézióban (az aljzathoz tapadnak); gliko-
calix. Az exopoliszacharidok baktériumok általi szintézisén kívül van egy másik mechanizmus is a képződésükre: az extracelluláris bakteriális enzimek diszacharidokra gyakorolt hatására. Ennek eredményeként dextránok és levánok képződnek.
A kapszula és a nyálka megvédi a baktériumokat a károsodástól és a kiszáradástól, mivel hidrofil lévén jól megköti a vizet és megakadályozza a makroorganizmus és a bakteriofágok védőfaktorainak hatását.
Flagella baktériumok határozzák meg a baktériumsejt mobilitását. A flagellák vékony filamentumok, amelyek a citoplazmatikus membránból származnak, és hosszabbak, mint maga a sejt (2.7. ábra). A flagella vastagsága 12-20 nm, hossza 3-15 µm. 3 részből állnak: egy spirálszálból, egy horogból és egy alaptestből, amely speciális korongokkal ellátott rudat tartalmaz (1 pár korong a gram-pozitív baktériumokban és 2 pár a gram-negatív baktériumokban). A zászlók a citoplazma membránjához és a sejtfalhoz korongokkal kapcsolódnak. Ez azt a hatást hozza létre, hogy egy villanymotor egy rúddal - egy rotorral - forgatja a flagellumot. A citoplazma membránján lévő protonpotenciál különbséget energiaforrásként használják. A rotációs mechanizmust a proton ATP szintetáz biztosítja. A flagellum forgási sebessége elérheti a 100 rps-t. Ha egy baktériumnak több flagellája van, azok szinkronban forogni kezdenek, egyetlen köteggé fonódnak össze, egyfajta légcsavart alkotva.
A flagella fehérjéből áll - flagellinből (a. flagellum - flagellum), amely egy antigén - az úgynevezett H-antigén. A flagellin alegységek spirálban vannak csavarodva.
A különböző fajokhoz tartozó baktériumok flagelláinak száma egytől (monotrichus) a Vibrio cholerae-ben a baktérium (peritrichus) kerülete mentén elterülő több tíz és száz flagelláig (Escherichia coliban, Proteusban stb.) változik. A Lophotrichusban egy köteg flagella található a sejt vége. Az Amphitriciának egy flagellum vagy egy köteg flagella van a sejt ellentétes végein.
A zászlók kimutatása nehézfémekkel permetezett készítmények elektronmikroszkópos vizsgálatával, vagy fénymikroszkóppal speciális, maratási és adszorpciós módszerekkel végzett kezelés után történik.
olyan anyagok, amelyek a flagellák vastagságának növekedéséhez vezetnek (például ezüstösödés után).
Villi, vagy ivott(fimbriae) - fonalszerű képződmények (2.7. ábra), vékonyabbak és rövidebbek (3 + 10 nm x 0,3 + 10 µm), mint a flagellák. A pilusok a sejtfelszínről nyúlnak ki, és a fehérje pilinből állnak. Antigén aktivitással rendelkeznek. Vannak pilusok, amelyek az adhézióért, azaz a baktériumok érintett sejthez való kötődéséért felelősek, valamint a táplálkozásért, a víz-só anyagcseréért és a szexuális (F-pili), vagy a konjugációs pilusokért.
Pili általában számos - több száz sejtenként. Azonban sejtenként általában 1-3 szexuális fűrésze van: ezeket az úgynevezett „férfi” donorsejtek alkotják, amelyek átvihető plazmidokat tartalmaznak. (F-, R-, Col plazmidok). Az ivarpilusok megkülönböztető jellemzője, hogy kölcsönhatásba lépnek speciális „férfi” gömb alakú bakteriofágokkal, amelyek intenzíven adszorbeálódnak a nemi pilusokon (2.7. ábra).
Vita- a nyugvó baktériumok sajátos formája, gram-pozitív típusú sejtfalszerkezettel (2.8. ábra).
A spórák a baktériumok létére kedvezőtlen körülmények között keletkeznek (száradás, tápanyaghiány stb.). A baktériumsejt belsejében egyetlen spóra (endospóra) képződik. A spórák képződése hozzájárul a faj megőrzéséhez, és nem szaporodási módszer, mint a gombáknál.
A nemzetség spóraképző baktériumai Bacilus, y amelyek spóramérete nem haladja meg a sejt átmérőjét, bacilusoknak nevezzük. A spóraképző baktériumokat, amelyekben a spóra mérete meghaladja a sejt átmérőjét, ezért orsó alakúak, clostridiumoknak nevezzük, például a nemzetségbe tartozó baktériumok. Clostridium (lat. Clostridium - orsó). A spórák savállóak, ezért Aujeszky-módszerrel vagy Ziehl-Neelsen-módszerrel vörösre festik, a vegetatív sejtet pedig kékre festik.
A spórák kialakulása, a spórák alakja és elhelyezkedése a sejtben (vegetatív) a baktériumok faji sajátossága, amely lehetővé teszi a baktériumok megkülönböztetését. A spórák alakja lehet ovális, gömb alakú; helye a sejtben terminális, azaz a rúd végén (a tetanusz kórokozójában), subterminálisan - közelebb a pálcika végéhez (a botulizmus kórokozóiban, gáz gangrénában) és központi az anthrax bacilusban. .
Folyamat sporuláció(sporuláció) egy sor szakaszon megy keresztül, melynek során a baktérium vegetatív sejt citoplazmájának és kromoszómájának egy része elválik, körülvéve növekvő citoplazmatikus membránnal - prospóra képződik. A prospórát két citoplazma membrán veszi körül, amelyek között vastag, módosított peptidoglikán kéregréteg (kéreg) képződik. Belülről érintkezik a spóra sejtfalával, kívülről pedig - a spóra belső héjával. A spóra külső héját vegetatív sejt alkotja. Egyes baktériumok spóráinak további burkolata van - exosporium. Ily módon többrétegű, rosszul áteresztő héj jön létre. A sporulációt a prospóra, majd a fejlődő spórahéj intenzív dipikolsav- és kalciumion-fogyasztása kíséri. A vita szerez hőellenállás, ami a kalcium-dipikolinát jelenlétével függ össze.
A spóra hosszú ideig fennmaradhat a többrétegű héj, a kalcium-dipikolinát, az alacsony víztartalom és a lassú anyagcsere-folyamatok miatt. A talajban például a lépfene és a tetanusz kórokozói évtizedekig fennmaradhatnak.
Kedvező körülmények között a spórák kicsíráznak, és három egymást követő szakaszon mennek keresztül:
motiváció, beavatás, növekedés. Ebben az esetben egy spórából egy baktérium képződik. Az aktiválás készen áll a csírázásra. 60-80 °C hőmérsékleten a spóra aktiválódik a csírázáshoz. A csírázás beindítása néhány percig tart. A kinövési szakaszt gyors növekedés jellemzi, amelyet a héj pusztulása és a palánta megjelenése kísér.

Tartalom Spanyol bogárból (vagy spanyol bogárból) nyert kivonat alapú étrend-kiegészítő...

Ekaterina MirimanovaSystem mínusz 60. RevolutionSystem mínusz 60 Ekaterina Mirimanovával"Rendszer mínusz 60. Revolution"...

Az elnehezülés és a puffadás mind a hétköznapi túlevés, mind a súlyosabb emésztőrendszeri problémák okai...

A második vércsoport, az Rh-negatív, sok évvel ezelőtt jelent meg, amikor az ember abbahagyta a vadászat, és...

AZ ANOREXIA JELENSÉG PSZICHOLÓGIAI VONATKOZÁSAI (KÍSÉRLETI TANULMÁNY) T. V. Tarasova, E. V. Arsentieva V...

Tartalom Mivel a bőr ezen a területen vékony, hajlamosabb a különböző típusú foltok megjelenésére. Vörösség alatt...

vseslav Szo, 2015.10.17. - 20:50 A Vasileostrovskaya állomás az egyik legrégebbi állomás...

Leggyakrabban a kecskegombát (Suillus bovinus) kecskegombának nevezik a gombászok köznyelvében. Ez a cső alakú...

Minden embernek voltak legalább egyszer álmai életében. Sokan nem tulajdonítanak nekik jelentőséget, de néhány...

Daedalus (kráter). Átmérő: 93 km Mélység: 3 km (NASA fotó) A Hold ősidők óta felkeltette az emberek figyelmét. A II...

Az elnökök napját február harmadik hétfőjén ünneplik az Egyesült Államokban. A 70-es évekig az ünnepeket a...

Az utolsó hívás napján csak egy szót szeretnék mondani szeretett osztályfőnökömnek: „Köszönöm.” De mennyi ez...

A görögdinnyét ízletes és egészséges bogyónak tartják, magvai különösen hasznosak. Kicsi, sötétbarna vagy...

„Beszédfejlődés 6 éves gyermekeknél” A gyermek nem születik fejlett beszéddel. Lehetetlen határozott választ adni arra a kérdésre, hogy...

Ekaterina MirimanovaSystem mínusz 60. RevolutionSystem mínusz 60 Ekaterina Mirimanovával„Rendszer mínusz 60....
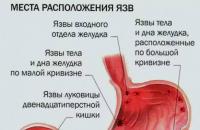
Az elnehezülés és a puffadás mind a hétköznapi túlevés, mind a komolyabb emésztési problémák okai...