Silkė po kailiu – klasikinis receptas
Kokie būtų Naujieji metai be šampano, mandarinų, Olivier, aspicų ir visų mėgstamo „Silkės po kailiu“. Su paskutiniu...
Spalva yra vizualinis, subjektyvus žmogaus suvokimas apie matomą šviesą, jos spektrinės sudėties skirtumus, jaučiamas akimis. Žmonių spalvų regėjimas yra daug geriau išvystytas nei kitų žinduolių.
Šviesa veikia šviesai jautrius tinklainės receptorius, o jie savo ruožtu sukuria signalą, kuris perduodamas į smegenis. Spalvos pojūtis, kaip ir visas daugiapakopis regėjimo suvokimas, kompleksiškai formuojasi grandinėje: akis (eksteroceptoriai ir tinklainės nerviniai tinklai) – regos smegenų sritis.
Šiuo atveju kūgiai yra atsakingi už spalvų suvokimą, strypai - už prieblandos matymą.Akys reaguoja į tris pagrindines spalvas: raudoną, žalią ir mėlyną. Žmogaus smegenys savo ruožtu suvokia spalvą kaip šių trijų signalų derinį. Jeigu susilpnėja arba išnyksta vienos iš trijų pagrindinių tinklainės spalvų suvokimas, tai žmogus tam tikros spalvos nesuvokia. Yra žmonių, kurie, pavyzdžiui, negali atskirti raudonos nuo žalios. Taigi apie septynis procentus vyrų ir apie pusę procento moterų kenčia nuo tokių problemų. Visiškas „spalvų aklumas“, kai receptorių ląstelės visiškai neveikia, yra labai retas. Kai kurie žmonės turi sunkumų dėl naktinio matymo, o tai paaiškinama silpnu lazdelių – jautriausių prieblandos regėjimo receptorių – jautrumu. Tai gali būti paveldimas veiksnys arba dėl vitamino A trūkumo. Tačiau žmogus prisitaiko prie „spalvų sutrikimų“, kurių be specialaus tyrimo beveik neįmanoma nustatyti. Normalaus regėjimo žmogus gali atskirti iki tūkstančio skirtingų spalvų.
Jautrumas ir regėjimo aštrumas. Jautrumas šviesos intensyvumui nustatomas pagal strypus ir kūgius. Tarp jų yra du reikšmingi skirtumai, paaiškinantys daugybę reiškinių, susijusių su intensyvumo arba ryškumo suvokimu.
Pirmasis skirtumas yra tas, kad vidutiniškai viena ganglinė ląstelė yra prijungta prie daugiau strypų nei kūgių; todėl „lazdelės“ ganglioninės ląstelės turi daugiau įėjimų nei „kūginės“ ląstelės. Antrasis skirtumas yra tas, kad strypai ir kūgiai yra skirtingai išsidėstę tinklainėje. Fovealinėje zonoje yra daug kūgių, bet nėra strypų, o periferijoje daug strypų, bet palyginti mažai kūgių. Kadangi ganglinė ląstelė yra sujungta su daugiau strypų nei kūgių, lazdelių matymas yra jautresnis nei kūgio regėjimas. Fig. 4.11 pav. tiksliai parodyta, kaip tai vyksta. Kairėje paveikslo pusėje yra trys gretimi kūgiai, kurių kiekvienas yra sujungtas (netiesiogiai) su viena gangliono ląstele; dešinėje paveikslėlio pusėje pavaizduoti trys gretimi strypai, kurie visi yra sujungti (ne tiesiogiai) su ta pačia gangliono ląstele.
Norėdami suprasti, ką reiškia šie skirtingi kūgio ir strypų „laidų“ raštai, įsivaizduokite, kad ant strypų ir kūgių yra trys labai silpnos, glaudžiai išdėstytos šviesos dėmės. Pristačius kūgius, kiekviena šviesos dėmė atskirai gali būti per silpna, kad atitinkamame receptoriuje sukeltų nervinį impulsą, todėl joks nervinis impulsas nepasieks ganglioninės ląstelės. Bet kai ant strypų pateikiamos tos pačios trys dėmės, šių trijų receptorių aktyvacija gali būti sujungta, o tada šios sumos pakaks nerviniam atsakui sukelti gangliono ląstelėje. Todėl kelių lazdelių prijungimas prie vienos ganglinės ląstelės užtikrina nervinio aktyvumo konvergenciją ir būtent dėl šios konvergencijos lazdelių matymas yra jautresnis nei kūgio regėjimas.
Tačiau šis jautrumo pranašumas kainuoja, ty mažesnis regėjimo aštrumas, palyginti su kūgio regėjimu (regėjimo aštrumas yra galimybė atskirti detales). Dar kartą pažvelkime į dvi diagramas pav. 4.10, bet dabar įsivaizduokite, kad trys gretimos šviesos dėmės yra gana ryškios. Pateikus kūgius, kiekviena dėmė sukels nervinį atsaką atitinkamame receptoriuje, o tai savo ruožtu sukels nervinius impulsus trijose skirtingose ganglioninėse ląstelėse; Į smegenis bus išsiųsti trys skirtingi pranešimai, o sistema galės sužinoti apie trijų skirtingų objektų egzistavimą. Jei šios trys gretimos šviesos dėmės pateikiamos ant strypų, visų trijų receptorių nervinis aktyvumas bus sujungtas ir perduotas į vieną gangliono ląstelę; todėl į smegenis bus nusiųstas tik vienas pranešimas, o sistema neturės galimybės sužinoti apie daugiau nei vieno objekto egzistavimą. Trumpai tariant, tai, kaip receptoriai jungiasi prie ganglioninių ląstelių, paaiškina lazdelės ir kūgio regėjimo jautrumo ir aštrumo skirtumus.
Kita šių skirtumų pasekmė yra ta, kad žmonės geriau aptinka silpną šviesą strypo periferijoje nei duobėje.
Taigi, nors regėjimo aštrumas fovea yra didesnis nei periferijoje, jautrumas yra didesnis periferijoje. Tai, kad jautrumas periferijoje yra didesnis, galima nustatyti išmatuojant absoliučią objekto slenkstį, kai jam blyksteli šviesa tamsiame kambaryje. Slenkstis bus žemesnis (tai reiškia didesnį jautrumą), jei subjektas žiūri šiek tiek į šoną, kad blyksniai būtų matomi esant periferiniam matymui, nei tada, jei jis žiūri į blyksnius tiesiogiai ir šviesa patenka į duobutę. Jau matėme vieną iš pasekmių, kai periferijoje bus mažiau kūgių (žr. 4.9 pav.). Strypų pasiskirstymo poveikį galima aptikti, kai žiūrime į žvaigždes naktį. Galbūt pastebėjote, kad norint kuo aiškiau matyti silpną žvaigždę, reikia šiek tiek pakeisti žiūrėjimo kryptį į vieną žvaigždės kraštą. Dėl šios priežasties žvaigždės šviesa suaktyvina maksimalų įmanomą strypų skaičių.
Šviesos pritaikymas. Iki šiol pabrėžėme, kad žmonės yra jautrūs stimuliacijos pokyčiams. Kita medalio pusė yra ta, kad jei stimulas nepasikeičia, žmogus prie jo prisitaiko. Gerą apšvietimo pritaikymo pavyzdį galima pamatyti įėjus į tamsų kino teatrą iš saulės apšviestos gatvės. Iš pradžių vos ką nors matote silpnoje šviesoje, atsispindėjusioje nuo ekrano. Tačiau po kelių minučių matai pakankamai gerai, kad rastum vietą. Po kurio laiko galite atskirti veidus esant silpnam apšvietimui. Vėl išėjus į ryškiai apšviestą gatvę iš pradžių beveik viskas atrodo skausmingai šviesu, o tokioje ryškioje šviesoje neįmanoma nieko atskirti. Tačiau viskas grįžta į normalią greičiau nei per minutę, nes greičiau prisitaikoma prie ryškesnės šviesos. Fig. 4.12 paveiksle parodyta, kaip tamsoje laikui bėgant absoliuti riba mažėja. Kreivė susideda iš dviejų šakų. Viršutinė šaka yra susijusi su kūgių darbu, o apatinė šaka - su strypais. Meškerykočių sistema prisitaiko daug ilgiau, tačiau yra jautri daug silpnesnei šviesai.

Žmogaus regėjimo ypatybės
Žmogus nemato visiškoje tamsoje. Kad žmogus matytų objektą, šviesa turi atsispindėti nuo objekto ir pataikyti į tinklainę. Šviesos šaltiniai gali būti natūralūs (ugnis, Saulė) ir dirbtiniai (įvairios lempos).
Žmogaus akis yra radijo imtuvas, galintis priimti tam tikro (optinio) dažnių diapazono elektromagnetines bangas. Pirminiai šių bangų šaltiniai yra jas skleidžiantys kūnai (saulė, lempos ir kt.), antriniai šaltiniai – kūnai, atspindintys pirminių šaltinių bangas. Šviesa iš šaltinių patenka į akis ir daro jas matomas žmonėms. Taigi, jei kūnas yra skaidrus bangoms matomo dažnio diapazone (oras, vanduo, stiklas ir kt.), tada akis jo aptikti negali.
Regėjimo dėka mes gauname 90% informacijos apie mus supantį pasaulį, todėl akis yra vienas svarbiausių jutimo organų. Akis gali būti vadinama sudėtingu optiniu įrenginiu. Pagrindinė jo užduotis yra „perduoti“ teisingą vaizdą į regos nervą.
Žmogaus akies jautrumas šviesai
Akies gebėjimas suvokti šviesą ir atpažinti įvairų jos ryškumo laipsnį vadinamas šviesos suvokimu, o gebėjimas prisitaikyti prie skirtingo apšvietimo ryškumo – akies prisitaikymu; šviesos jautrumas vertinamas pagal šviesos dirgiklio slenkstinę vertę. Žmogus, turintis gerą regėjimą, naktį gali matyti žvakės šviesą kelių kilometrų atstumu. Maksimalus šviesos jautrumas pasiekiamas po pakankamai ilgo prisitaikymo prie tamsos.
Žmogaus akyje yra dviejų tipų šviesai jautrių ląstelių (receptorių): labai jautrūs strypai, atsakingi už prieblandos (naktinį) matymą, ir mažiau jautrūs kūgiai, atsakingi už spalvų matymą.
Žmogaus tinklainėje yra trijų tipų kūgiai, kurių didžiausias jautrumas pasireiškia raudonoje, žalioje ir mėlynoje spektro dalyse. Kūgių tipų pasiskirstymas tinklainėje yra netolygus: „mėlyni“ kūgiai randami arčiau periferijos, o „raudoni“ ir „žali“ kūgiai pasiskirstę atsitiktinai. Kūgio tipų atitikimas trims „pagrindinėms“ spalvoms leidžia atpažinti tūkstančius spalvų ir atspalvių. Trijų tipų kūgių spektrinio jautrumo kreivės iš dalies sutampa, o tai prisideda prie metamerizmo reiškinio. Labai stipri šviesa sužadina visų 3 tipų receptorius, todėl suvokiama kaip akinanti baltoji spinduliuotė.
Vienodas visų trijų elementų stimuliavimas, atitinkantis svertinį dienos šviesos vidurkį, taip pat sukuria baltos spalvos pojūtį. Žmogaus spalvų regėjimą kontroliuoja genai, koduojantys šviesai jautrius opsino baltymus. Pasak trijų komponentų teorijos šalininkų, spalvų suvokimui pakanka trijų skirtingų baltymų, kurie reaguoja į skirtingus bangos ilgius. Dauguma žinduolių turi tik du iš šių genų, todėl jie turi juodą ir baltą regėjimą.
Žmogus mato ne akimis, o akimis, iš kur informacija per regos nervą, chiazmą, regos takus perduodama į tam tikras smegenų žievės pakaušio skilčių sritis, kur matomas išorinio pasaulio vaizdas. susiformavo. Visi šie organai sudaro mūsų regos analizatorių arba regos sistemą.
Regėjimo pokyčiai su amžiumi
Naujagimiams ir ikimokyklinio amžiaus vaikams lęšiukas yra labiau išgaubtas ir elastingesnis nei suaugusiojo, jo laužiamoji galia didesnė. Tai leidžia vaikui aiškiai matyti objektą, esantį mažesniu atstumu nuo akies nei suaugęs. Ir jei kūdikiui jis yra skaidrus ir bespalvis, suaugusiojo lęšis turi šiek tiek gelsvą atspalvį, kurio intensyvumas gali didėti su amžiumi. Tai neturi įtakos regėjimo aštrumui, bet gali turėti įtakos mėlynos ir violetinės spalvos suvokimui. Sensorinės ir motorinės regos funkcijos vystosi vienu metu. Pirmosiomis dienomis po gimimo akių judesiai yra asinchroniniai, kai viena akis nejuda, galima stebėti kitos. Gebėjimas fiksuoti daiktą žvilgsniu susiformuoja nuo 5 dienų iki 3–5 mėnesių amžiaus. Reakcija į daikto formą jau pastebima 5 mėnesių vaikui. Ikimokyklinio amžiaus vaikams pirmą reakciją sukelia daikto forma, vėliau jo dydis ir galiausiai spalva. Regėjimo aštrumas didėja su amžiumi, gerėja ir stereoskopinis regėjimas. Stereoskopinis regėjimas(iš graikų στερεός - kietas, erdvinis) - regėjimo tipas, kuriame galima suvokti objekto formą, dydį ir atstumą, pavyzdžiui, žiūronų regėjimo dėka Stereoskopinis regėjimas pasiekia optimalų lygį iki 17 metų amžiaus. –22 m., o nuo 6 metų mergaičių stereoskopinis regėjimo aštrumas yra didesnis nei berniukų. Matymo laukas sparčiai didėja. Iki 7 metų jo dydis yra maždaug 80% suaugusiojo regėjimo lauko dydžio. Po 40 metų sumažėja periferinio matymo lygis, tai yra, susiaurėja regėjimo laukas ir pablogėja vaizdas iš šono. Maždaug po 50 metų ašarų skysčio gamyba mažėja, todėl akys mažiau hidratuojamos nei jaunesniame amžiuje. Pernelyg didelis sausumas gali pasireikšti akių paraudimu, skausmu, ašarojančiomis akimis veikiant vėjui ar ryškiai šviesai. Tai gali priklausyti nuo įprastų veiksnių (dažno akių įtempimo ar oro taršos). Su amžiumi žmogaus akis ima blankiau suvokti aplinką, mažėja kontrastas ir ryškumas. Gebėjimas atpažinti spalvas, ypač artimas spalvas, taip pat gali būti sutrikęs. Tai tiesiogiai susiję su tinklainės ląstelių, kurios suvokia spalvų atspalvius, kontrastą ir ryškumą, skaičiaus sumažėjimą. Kai kuriuos su amžiumi susijusius regėjimo sutrikimus sukelia presbiopija, kuri pasireiškia neaiškiais, neryškiais vaizdais bandant žiūrėti į arti akių esančius objektus. Gebėjimas sufokusuoti regėjimą į mažus objektus reikalauja maždaug 20 dioptrijų (fokusuojant į objektą, esantį 50 mm atstumu nuo stebėtojo) vaikams, iki 10 dioptrijų 25 metų amžiaus (100 mm) ir 0,5–1 dioptrijų 60 metų amžiaus ( gebėjimas sutelkti dėmesį į objektą, esantį už 1–2 metrų). Manoma, kad taip nutinka dėl vyzdį reguliuojančių raumenų susilpnėjimo, pablogėja ir vyzdžių reakcija į į akį patenkantį šviesos srautą. Todėl kyla sunkumų skaitant esant silpnam apšvietimui, o prisitaikymo laikas pailgėja, kai keičiasi apšvietimas.
Taip pat su amžiumi greičiau pradeda atsirasti regos nuovargis ir net galvos skausmai.
Spalvų suvokimo psichologija
Spalvų suvokimo psichologija – žmogaus gebėjimas suvokti, atpažinti ir įvardyti spalvas. Spalvos suvokimas priklauso nuo fiziologinių, psichologinių, kultūrinių ir socialinių veiksnių komplekso. Iš pradžių spalvų suvokimo tyrimai buvo vykdomi spalvų mokslo rėmuose; Vėliau prie problemos prisijungė etnografai, sociologai ir psichologai. Regėjimo receptoriai teisėtai laikomi „smegenų dalimi, iškelta į kūno paviršių“. Nesąmoningas regėjimo suvokimo apdorojimas ir korekcija užtikrina regėjimo „teisingumą“, o tai yra ir „klaidų“ priežastis vertinant spalvą tam tikromis sąlygomis. Taigi, pašalinus „foninį“ akies apšvietimą (pavyzdžiui, žiūrint į tolimus objektus per siaurą vamzdelį), žymiai pakeičiamas šių objektų spalvos suvokimas. Dėl akies prigimties šviesa, sukelianti tos pačios spalvos pojūtį (pavyzdžiui, baltą), tai yra vienodą trijų regos receptorių sužadinimo laipsnį, gali turėti skirtingą spektrinę sudėtį. Daugeliu atvejų žmogus šio efekto nepastebi, tarsi „atspėdamas“ spalvą. Taip yra todėl, kad nors skirtingo apšvietimo spalvinė temperatūra gali būti vienoda, tačiau to paties pigmento atspindimos natūralios ir dirbtinės šviesos spektrai gali labai skirtis ir sukelti skirtingą spalvų pojūtį.
Periferinis regėjimas(laukas regėjimas) – apibrėžkite lauko ribas regėjimas projektuojant juos ant sferinio paviršiaus (naudojant perimetrą).
Čia pažvelgsime į kai kuriuos mokslinius duomenis iš fizikos ir fiziologijos, kad suprastume, kaip vyksta suvokimo procesas.
Pradėkime nuo vaizdo kanalo. Vizija yra informatyviausias informacijos kanalas. Per jį gauname didžiausią informacijos kiekį iš išorinio pasaulio. Iš fizikos žinome, kad regėjimas yra šviesos iš aplinkos suvokimas. Didžiausias šviesos šaltinis Žemėje yra Saulė. Šviesa pagal savo prigimtį yra tam tikro dažnio elektromagnetinė banga.
Subjektyviai šias bangas suvokiame kaip tam tikrą spalvą. Pavyzdžiui, šviesą, kurios dažnis yra 400-480 THz, mes suvokiame kaip raudoną, o šviesą, kurios dažnis yra 620-680 THz, kaip mėlyną. Šiek tiek vėliau aptarsime, kodėl mes taip suvokiame šiuos šviesos dažnius. Tiesą sakant, jei paimtume visą elektromagnetinės spinduliuotės dažnių diapazoną, pamatytume, kad matomą šviesą suvokiame tik labai trumpą dažnių diapazoną. Likusio mes nesuvokiame, t.y. yra banga, bet mes jos nematome. Pavyzdžiui, nematome radijo bangų, kurias priima jūsų televizorius, nors jos fiziškai yra erdvėje.
Šviesos spindulys, sklindantis iš saulės, turi visą pluoštą skirtingo dažnio elektromagnetinių bangų. Iš esmės šiame šviesos pluošte yra beveik visų dažnių bangų. Šis šviesos spindulys vadinamas balta šviesa. Norėdami pamatyti, kad baltoje šviesoje yra visų dažnių bangos, tereikia nukreipti tą šviesos spindulį į prizmę ir tai matome.
Balta šviesa suskilo į visų spalvų vaivorykštę. Atrodė, kad prizmė atskyrė skirtingo dažnio bangas skirtingomis kryptimis.
Dabar pažiūrėkime, kaip paaiškėja, kad mus supantys objektai turi skirtingas spalvas. Ant objekto krentant baltam šviesos pluoštui, objekto paviršius sugeria beveik visas skirtingo dažnio bangas ir atspindi tam tikro siauro dažnių diapazono bangas. Jei, pavyzdžiui, baltas šviesos spindulys nukrenta ant raudono objekto paviršiaus, tai šis objektas pats sugers visas bangas, kurių dažnis skiriasi nuo raudonos dažnio, o bangos, kurių dažnis yra raudonas, atsispindės nuo jo paviršiaus.
Atminkite, kad sakydamas „raudonas dažnis“ neturiu omenyje, kad banga iš tikrųjų yra raudona. Tai reiškia, kad šios bangos dažnis yra 400–480 THz diapazone. Ne daugiau. Pati šviesos banga neturi jokių spalvų.
Taigi, raudono dažnio šviesos banga atsispindi nuo objekto įvairiomis kryptimis. Tada ši nuo objekto atsispindėjusi šviesa patenka į mūsų akis. Skirtingi objektai mums atrodo skirtingomis spalvomis, nes šių objektų paviršiai skirtingai atspindi ant jų krentančią baltą šviesą. Vieni atspindi daugiausia raudonas bangas, kiti – žalias, kiti sugeria beveik visas bangas, tada objektas mums atrodo juodas.
Kas nutinka, kai į akis patenka skirtingų dažnių šviesa? Akių tinklainėje yra šviesos receptoriai – kūgiai ir lazdelės. Yra trijų tipų kūgiai: vieni geriausiai šviesą suvokia mėlynai violetinėje srityje, kiti – geltonai žalioje, kiti – raudonoje. Tie. skirtingi kūgiai reaguoja į šviesos bangas iš tam tikro dažnių diapazono.

Tada tinklainės kūgiai sukuria nervinį impulsą. Šis impulsas keliauja iš tinklainės nervų skaidulomis (neuronais) į žmogaus smegenis. Žmogaus smegenyse yra sritis, kuri apdoroja iš akių gaunamus signalus. smegenų regėjimo sritis. Pačios smegenys yra didžiulė kolekcija neuronai. Tai ląstelės, susidedančios iš kūno, vieno aksono ir tūkstančių dendritų.

Dendritai yra neurono procesai, kurie gauna signalą iš kito neurono aksono. Aksonas yra neurono tęsinys, perduodantis signalą iš to neurono į kitus neuronus. Be to, aksonas šakojasi gale ir todėl gali perduoti signalą iš tam tikro neurono į kelis neuronus vienu metu.
Visi smegenų neuronai yra sujungti vienas su kitu per aksonus ir dendritus. Tūkstančiai neuronų jungiasi prie vieno neurono per dendritus ir per savo aksonus perduoda jam signalus. Tada neuronas apibendrina visus signalus į vieną ir per savo aksoną perduoda jį kitiems neuronams, su kuriais jis yra prijungtas. Rezultatas yra savotiškas neuroninis tinklas, jungiantis milijardus smegenų ląstelių.

Be neuronų, smegenyse taip pat yra vadinamųjų glijos ląstelės. Jie atlieka papildomas funkcijas ir tarnauja neuronams užtikrinant signalo perdavimą. Iš esmės smegenyse nėra nieko kito.
Taigi, signalas iš akies patenka į smegenų regėjimo zoną, esančią pakaušyje. Be to, iš regėjimo zonos signalas išsišakoja ir patenka į kitas smegenų dalis, įskaitant smegenų žievę, kur signalai paverčiami vaizdiniais, kuriuos mes suvokiame.
Noriu pabrėžti, kad niekur smegenyse nėra paveikslėlių. Viskas, kas yra, yra tik nerviniai impulsai, pereinantys iš vieno neurono į kitą.
Smegenys skiria skirtingo diapazono šviesos bangas tik tuo, kad skirtingi kūgiai reaguoja į skirtingus šviesos bangų dažnius. Tada iš šių kūgių gaunamas įprastas elektrinis signalas. Smegenų regėjimo sritis išskiria spalvas pagal tai, iš kurių kūgių gaunamas signalas. Pats signalas neturi spalvos.
Atrodo, kad taip veikia regėjimas. Šviesa, kaip ir skirtingo dažnio elektromagnetinės bangos, atsispindi nuo objektų ir patenka į mūsų akis. Daiktų paviršius dalį bangų sugeria, dalį atspindi (tai priklauso nuo paviršiaus savybių). Atsispindinčios bangos patenka į mūsų akis, kur tinklainėje esančių kūgių ir strypų pagalba paverčiamos nerviniais impulsais. Šie nerviniai impulsai neuronų tinklu keliauja į smegenis, tiksliau į smegenų regėjimo sritį. Iš regėjimo srities signalas plinta į kitas smegenų dalis. Be neuronų tinklo, palaikančio glijos ląsteles ir nervinius signalus, smegenyse nėra nieko kito.
Dabar trumpai apsvarstykime likusių suvokimo kanalų veikimą. Šios suvokimo kanalų veikimo schemos iš esmės nesiskiria nuo vizualinio kanalo veikimo schemos.
Garsas pagal savo prigimtį yra oro vibracija. Tie. objektas dėl to, kad jis vibruoja, sukelia vibracijas aplink jį esančiame ore. Šios vibracijos sklinda oru įvairiomis kryptimis ir galiausiai patenka į žmogaus ausis. Jei nebūtų oro, objektas neperduotų virpesių ir nebūtų garso.
Garso bangos, kaip ir šviesos bangos, turi skirtingą dažnį. Kuo mažesnis garso bangos virpesių dažnis, tuo subjektyviai mums atrodo, kad garsas yra žemesnis. Tai taikoma bosui. Kuo aukštesni garso bangos virpesiai, tuo subjektyviai mums atrodo, kad garsas yra aukštesnis ir girgždantis.
Tačiau aukštis neturi nieko bendra su garso bangomis. Garso bangos yra tiesiog skirtingo dažnio bangos, sklindančios oru. Pačios šios bangos neturi jokio garso.
Tada objektų garso bangos pasiekia mūsų ausis. Ausyje yra būgnelis, kuris subtiliai reaguoja į oro, patenkančio į ausį, virpesius. Jis vibruoja tokiu pat dažniu kaip ir garso banga, patenkanti į ausį. Toliau, naudojant sudėtingą ausies virpesių konvertavimo sistemą, garso banga paverčiama nerviniu impulsu, kuris klausos nervu keliauja į smegenis, į tuos skyrius, kurie yra atsakingi už klausos informacijos apdorojimą.

Taigi, kaip ir šviesa, garsas taip pat paverčiamas nerviniu impulsu, kurį apdoroja smegenys. Nervinis impulsas, kuris ateina iš akių, nesiskiria nuo nervinio impulso, kuris ateina iš ausų. Smegenyse vyksta visa šių signalų diskriminacija ir nustatymas, kokį signalą jie neša. Smegenys nustato tai, per kokius nervų kelius signalas atėjo. Jei nervinis impulsas (signalas) atkeliavo iš neuronų, atsakingų už šviesos suvokimą, smegenys šį signalą interpretuos kaip vaizdinį. Jei signalas ateina iš neuronų, atsakingų už garso suvokimą, smegenys šį signalą interpretuos kaip garsinį (garsą).
Kalbant apie lytėjimą, kvapą ir skonį, galime trumpai pasakyti štai ką. Oda turi specialius receptorius, kurie reaguoja į prisilietimą ir oro temperatūrą. Toliau viskas vyksta pagal tą pačią schemą. Šių receptorių nervinis signalas pasiekia smegenis.
Nosyje yra receptorių, kurie reaguoja į tam tikras molekules. Pavyzdžiui, rožės žiedas išskiria molekules. Šios molekulės patenka į nosį, o uoslės receptoriai reaguoja į tam tikras molekules. Tada uoslės receptoriai perduoda signalą į smegenis.
Kalbant apie skonį, liežuvyje yra atitinkami receptoriai, kurie reaguoja į medžiagų molekules, kurios patenka į žmogaus burną. Ir vis tiek pagal schemą nerviniai signalai iš šių receptorių patenka į smegenis.
Taigi, atkreipsiu jūsų dėmesį į tai, kad išorinis pasaulis neneša jokių paveikslų, garsų, skonių ar pojūčių. Viskas, kas egzistuoja išoriniame pasaulyje, yra įvairios bangos ir medžiagų molekulės. A tai, ką matome, girdime ir jaučiame, yra mūsų smegenų darbo rezultatas. Atėjo laikas užduoti svarbų klausimą: kodėl signalai iš regos smegenų srities suvokiami būtent taip, kaip mes juos suvokiame, t.y. trimačio paveikslo pavidalu? Kodėl signalai iš smegenų srities, atsakingos už garsą, suvokiami kaip garsas? Juk nei šviesos bangose, nei oro virpesiuose nėra tokių savybių kaip spalva ir garsas.
Straipsnyje pateikiami duomenys apie aukštesniųjų gyvūnų ir žmonių regėjimo ciklo funkcionavimą. Chromoforo tinklainės turinčio transmembraninio receptoriaus baltymo rodopsino, atsakingo už šviesos suvokimo funkcijas, kai ją sugeria šviesos kvantinė molekulė, ir vėlesnes biochemines reakcijas, susijusias su katijonų (Na + /Ca 2+) kanalų uždarymu, fotociklas. ir membranos hiperpoliarizacija. Parodytas rodopsino sąveikos su receptoriumi G baltymo transducinu mechanizmas, kuris yra pagrindinis biocheminis regėjimo proceso žingsnis, susidedantis iš transducino aktyvinimo jo sąveikos su aktyvuotu rodopsinu metu ir surišto GTP keitimo į HDP. . Tada kompleksas disocijuoja ir aktyvuoja fosfodiesterazę, pakeisdamas jos slopinamąjį subvienetą. Taip pat atsižvelgiama į spalvų suvokimo mechanizmą vizualiniu aparatu, kuris turi galimybę analizuoti tam tikrus optinio spektro diapazonus kaip spalvas. Sumaišius žalią ir raudoną, neatsiranda vidurinė spalva: smegenys ją suvokia kaip geltoną. Skleisdamos elektromagnetines bangas, atitinkančias žalią ir raudoną, smegenys suvokia „vidurinį tirpalą“ - geltoną.
Vizija (vizualinis suvokimas) yra psichofiziologinio supančio pasaulio objektų vaizdų apdorojimo procesas, kurį atlieka regėjimo sistema ir leidžia susidaryti vaizdą apie aplinkinių objektų dydį, formą ir spalvą, jų santykinę padėtį. ir atstumas tarp jų. Per regėjimą žmogus gauna 90% visos informacijos, patenkančios į smegenis. Neatsitiktinai regėjimo vaidmuo žmogaus gyvenime yra toks milžiniškas. Regėjimo pagalba žmogus ne tik gaus didžiulį kiekį informacijos apie išorinį pasaulį, bet ir galės mėgautis gamtos grožiu bei puikiais meno kūriniais. Vizualinio suvokimo šaltinis yra išorinio pasaulio objektų skleidžiama arba atspindima šviesa.
Regėjimo funkcija atliekama dėl sudėtingos įvairių tarpusavyje susijusių struktūrų sistemos – regos analizatoriaus, susidedančio iš periferinės dalies (tinklainės, regos nervo, regos trakto) ir centrinės dalies, jungiančios vidurinių smegenų subkortikinius ir kamieninius centrus, taip pat smegenų žievės regėjimo sritis. Žmogaus akis suvokia tik tam tikro ilgio šviesos bangas – nuo 380 iki 770 nm. Šviesos spinduliai iš aptariamų objektų praeina per akies optinę sistemą (rageną, lęšį ir stiklakūnį) ir patenka į tinklainę, kurioje yra šviesai jautrios ląstelės – fotoreceptoriai (kūgiai ir strypai). Šviesa, patekusi į fotoreceptorius, sukelia juose esančių regos pigmentų (ypač labiausiai ištirto rodopsino, kuris yra atsakingas už elektromagnetinės spinduliuotės suvokimą matomame diapazone) biocheminių reakcijų, o tai savo ruožtu sukelia nerviniai impulsai, kurie perduodami į sekančius tinklainės neuronus ir toliau į regos nervą. Išilgai regos nervų, tada išilgai regėjimo takų nerviniai impulsai patenka į šoninį geniculate kūną - subkortikinį regėjimo centrą, o iš jo į žievės regėjimo centrą, esantį pakaušio smegenų skiltyse, kur susidaro regėjimas. atsiranda vaizdas.
Per pastarąjį dešimtmetį Rusijos ir užsienio mokslininkai gavo naujų duomenų, atskleidžiančių vizualinio suvokimo molekulinį pagrindą. Buvo nustatytos vaizdinės molekulės, dalyvaujančios reakcijoje į šviesą, ir atskleistas jų veikimo mechanizmas. Šiame straipsnyje nagrinėjami pagrindiniai biocheminiai mechanizmai, susiję su vizualiniu suvokimu ir vizualinių molekulių evoliucija.
Šviesos suvokimo procesas turi specifinę lokalizaciją tinklainės fotoreceptorinėse ląstelėse, kurios yra jautrios šviesai. Tinklainė yra daugiasluoksnis šviesai jautraus nervinio audinio sluoksnis, kuris iškloja vidinę akies obuolio užpakalinę dalį. Tinklainė yra ant pigmentinės membranos, vadinamos tinklainės pigmentiniu epiteliu (RPE), kuri sugeria šviesą, praeinančią per tinklainę. Tai neleidžia šviesai atsispindėti atgal per tinklainę ir vėl reaguoti, o tai neleidžia regėjimui susilieti.
Šviesa prasiskverbia į akį ir sukuria sudėtingą biocheminę reakciją šviesai jautriose tinklainės fotoreceptorių ląstelėse. Fotoreceptorių ląstelės skirstomos į dvi rūšis, kurios dėl būdingos formos vadinamos lazdelėmis ir kūgiais (1 pav.). Strypai yra spalvotame tinklainės sluoksnyje, kuriame sintetinamas fotochrominis baltymas rodopsinas, atsakingas už spalvų suvokimą, ir yra mažo intensyvumo šviesos receptoriai. Kūgiai išskiria grupę regėjimo pigmentų (jodopsino) ir yra pritaikyti spalvoms atskirti. Strypai leidžia matyti nespalvotus vaizdus prietemoje; Kūgiai suteikia spalvų matymą ryškioje šviesoje. Žmogaus tinklainėje yra apie 3 milijonai kūgių ir 100 milijonų lazdelių. Jų matmenys labai maži: ilgis apie 50 mikronų, skersmuo – nuo 1 iki 4 mikronų.
Kūgių ir strypų generuojamus elektrinius signalus apdoroja kitos tinklainės ląstelės – bipolinės ląstelės ir ganglioninės ląstelės – prieš perduodant juos į smegenis per regos nervą. Be to, yra dar du tarpinių neuronų sluoksniai. Horizontalios ląstelės perduoda pranešimus pirmyn ir atgal tarp fotoreceptorių ląstelių, bipolinių ląstelių ir viena kitos. Aamakrino ląstelės (tinklainės ląstelės) yra tarpusavyje susijusios su bipolinėmis ląstelėmis, ganglioninėmis ląstelėmis, taip pat viena su kita. Abu tokių interneuronų tipai atlieka pagrindinį vaidmenį apdorojant vaizdinę informaciją tinklainės lygmenyje, prieš perduodant ją į smegenis galutiniam apdorojimui.
Kūgiai yra maždaug 100 kartų mažiau jautrūs šviesai nei strypai, tačiau daug geriau atpažįsta greitus judesius. Strypą gali sužadinti vienas fotonas, mažiausias įmanomas šviesos kiekis. Molekulinių sąveikų kaskados sustiprina šį informacijos „kvantą“ į cheminį signalą, kurį vėliau suvokia nervų sistema. Signalo stiprinimo laipsnis skiriasi priklausomai nuo foninio apšvietimo: strypai yra jautresni prietemoje nei ryškioje šviesoje. Dėl to jie efektyviai veikia esant įvairiam foniniam apšvietimui. Strypų jutimo sistema supakuota į aiškiai atskiriamas ląstelines struktūras, kurias galima lengvai izoliuoti ir ištirti. in vitro.
Kūgiai ir strypai yra panašios struktūros ir susideda iš keturių dalių. Jų struktūroje įprasta išskirti:
išorinis segmentas, kuriame yra membraniniai pusdiski;
vidinis segmentas, kuriame yra mitochondrijos;
jungiamoji sekcija - susiaurėjimas;
sinaptinė sritis.
Strypo struktūra yra ilga plona ląstelė, padalinta į dvi dalis. Išoriniame ląstelės segmente yra dauguma molekulinių mechanizmų, kurie aptinka šviesą ir inicijuoja nervinius impulsus. Vidinis segmentas yra atsakingas už energijos generavimą ir molekulių atnaujinimą išoriniame segmente. Be to, vidinis segmentas sudaro sinapsinį terminalą, kuris palaiko ryšį su kitomis ląstelėmis. Jei izoliuota tinklainė šiek tiek papurtoma, išoriniai strypų segmentai nukrenta ir galima ištirti visą sužadinimo aparatą. in vitro labai išgryninta forma. Ši strypų savybė daro juos nepakeičiamu biochemikų tyrimo objektu.
Išorinis strypo segmentas yra siauras vamzdis, užpildytas plonų membraninių diskų krūva; suformuotas citoplazminės membranos ir nuo jos atskirtas. Vienoje kameroje jų yra apie 2 tūkst. Tiek vamzdelį, tiek diskus sudaro to paties tipo dviejų sluoksnių citoplazminė membrana. Tačiau lazdelės išorinė (plazminė) membrana ir diskų membrana atlieka skirtingas funkcijas, fotorecepciją ir nervinių impulsų generavimą. Diskuose yra dauguma baltymų molekulių, dalyvaujančių šviesos sugertyje ir sužadinimo atsako inicijavimu. Išorinė membrana skirta cheminį signalą paversti elektriniu.
Ryšys tarp dviejų segmentų atliekamas per citoplazmą ir blakstienų porą, pereinančią iš vieno segmento į kitą. Blakstienos turi tik 9 periferinius mikrovamzdelių dubletus: blakstienoms būdingos centrinių mikrotubulių poros nėra. Vidinis strypo segmentas yra aktyvios metabolizmo sritis; jis užpildytas mitochondrijomis, aprūpinančiomis energiją regėjimo procesams, ir poliribosomų, ant kurių sintetinami baltymai, dalyvaujantys membraninių diskų formavime ir regos pigmentas rodopsinas.
Viena iš svarbiausių integruotų transmembraninių receptorių G baltymų molekulių, susijusių su disko membrana, yra rodopsinas. Tai yra lazdelės fotoreceptoriaus chromoforo baltymas, kuris sugeria fotoną ir sukelia atsaką – tai pirmasis žingsnis įvykių grandinėje, kuri sukuria regėjimą. Rodopsiną sudaro du komponentai – bespalvis opsino baltymas, kuris veikia kaip fermentas, ir kovalentiškai surištas chromoforo komponentas – vitamino A darinys, 11- cis-tinklainė, kuri priima šviesą (2 pav.). Šviesos fotono sugertis 11- cis- tinklainė „įjungia“ fermentinį opsino aktyvumą ir suaktyvina biocheminę šviesai jautrių reakcijų kaskadą, atsakingą už regimąjį suvokimą.

Rodopsinas priklauso G-receptorių (GPCR receptorių) šeimai, atsakingai už transmembraninio signalo perdavimo mechanizmą, pagrįstą sąveika su tarpląstelinės membranos G-baltymais – signalizuojančiais G-baltymais, kurie yra universalūs tarpininkai perduodant hormoninius signalus iš ląstelės. membranos receptorius į efektorinius baltymus, sukeldami galutinį ląstelių atsaką. Jo erdvinės struktūros nustatymas yra svarbus biologijoje ir medicinoje, nes rodopsinas, kaip GPCR receptorių šeimos „protėvis“, yra daugelio kitų receptorių struktūros ir funkcijų „modelis“, kurie yra nepaprastai svarbūs moksliniu, fundamentaliu ir praktiniu požiūriu. (farmakologiniai) požiūriai.
Erdvinė rodopsino struktūra ilgą laiką negalėjo būti ištirta „tiesioginiais“ metodais – rentgeno difrakcine analize ir BMR spektroskopija, o kito transmembraninio baltymo bakteriorodopsino, giminingo rodopsinui, molekulinės struktūros panašios struktūros, atliekančio funkcijas. Nuo ATP priklausomos translokazės halofilinių mikroorganizmų ląstelių membranose, pumpuojančios protonus per citoplazminę ląstelės membraną ir dalyvaujančios anaerobiniame fotosintezės fosforilinime (sintezėje be chlorofilo), buvo identifikuotas dar 1990 m. Vaizdinio rodopsino struktūra liko nežinoma iki 2003 m.
Pagal savo struktūrą opsino molekulė yra 348 aminorūgščių liekanų polipeptidinė grandinė. Opsino aminorūgščių seką nustatė Rusijos mokslininkai Yu.A. Ovčinikovo vardu pavadintame Bioorganinės chemijos institute. MM. Šemjakinas Maskvoje. Šie tyrimai suteikia svarbios informacijos apie šio svarbaus disko membraną apimančio baltymo trimatę struktūrą. Opsino polipeptidinė grandinė sudaro septynias transmembranines α-spiralines sritis, esančias per membraną ir sujungtas trumpomis nespiralinėmis sritimis. Kuriame N-galas yra tarpląsteliniame regione, ir C-α-spiralės galas - citoplazmoje. 11-oji molekulė yra susijusi su viena iš α-spiralių. cis-tinklainė, esanti netoli membranos vidurio, kad jos ilgoji ašis būtų lygiagreti membranos paviršiui (3 pav.). 11 lokalizacijos vieta cis-tinklainė, sujungta aldamino jungtimi su Lys-296 liekanos ε-amino grupe, esančia septintoje α-spirale. Taigi 11- cis-Tinklainė yra įterpta į sudėtingos, labai organizuotos baltymų aplinkos centrą lazdelės ląstelės membranoje. Ši aplinka suteikia fotocheminį tinklainės „derinimą“, paveikdama jos absorbcijos spektrą. Pati nemokama 11- cis-Tinklainė ištirpusioje formoje turi absorbcijos maksimumą ultravioletinėje spektro srityje - esant 380 nm bangos ilgiui, o rodopsinas sugeria žalią šviesą esant 500 nm. Šis šviesos bangos ilgio poslinkis yra svarbus funkciniu požiūriu: jis suderina rodopsino sugerties spektrą su į akį patenkančios šviesos spektru.

Rodopsino absorbcijos spektrą lemia chromoforo – liekanos 11- savybės. cis- tinklainė ir opsinas. Šis spektras stuburiniuose gyvūnuose turi du maksimumus – vieną ultravioletinėje srityje (278 nm), dėl opsino, ir kitą matomoje srityje (apie 500 nm) – chromoforo sugertį (4 pav.). Vaizdo pigmento transformacija veikiant šviesai į galutinį stabilų produktą susideda iš daugybės labai greitų tarpinių etapų. Ištyrus rodopsino ekstraktuose esančių tarpinių produktų sugerties spektrus žemoje temperatūroje, kurioje šie produktai yra stabilūs, pavyko detaliai aprašyti visą vizualinio pigmento balinimo fotoprocesą.

Kai absorbuojamas molekulės 11- cis- tinklainės šviesos fotonas, jo molekulė izomerizuojasi į 11- visi- transas-tinklainė (kvantinė išeiga 0,67), o pats rodopsinas pakeičia spalvą (fotolizė). Šiuo atveju sukimasis vyksta aplink ryšį tarp 11 ir 12 anglies atomų cis-tinklainė, dėl kurios pasikeičia molekulės geometrija ir susidaro izomerinė forma - visi- transas-tinklainė be lenkimo, o po 10 ms įvyksta alosterinis rodopsino perėjimas į jo aktyvią formą (5 pav.). Sugerto šviesos fotono energija ištiesina grandinės vingį tarp 11 ir 12 anglies atomų. Šioje formoje 11- cis- tinklainė egzistuoja tamsoje. Stuburiniams gyvūnams rodopsino fotolizė baigiasi chromoforo atskyrimu nuo opsino; bestuburiuose chromoforas lieka surištas su baltymu visose fotolizės stadijose. Stuburiniuose gyvūnuose rodopsinas paprastai atsinaujina dėl opsino sąveikos su 11- cis-tinklainė, bestuburiuose - sugerus antrąjį šviesos fotoną.

Rodopsino molekulė, įterpta į lazdelės membraną, yra labai jautri šviesai (6 pav.). Nustatyta, kad šviesos fotono sugertis molekulėje pusėje atvejų sukelia 11-11 izomerizaciją. cis- tinklainė. Spontaniška tinklainės molekulės izomerizacija tamsoje vyksta labai retai – maždaug kartą per 1000 metų. Šis skirtumas turi svarbių pasekmių regėjimui. Kai vienas fotonas patenka į tinklainę, jį sugerianti rodopsino molekulė su ja labai efektyviai reaguoja, o milijonai kitų tinklainėje esančių rodopsino molekulių lieka „tyli“.

Vėlesni rodopsino fotocheminės transformacijos ir jo aktyvavimo ciklai sukelia regos nervo sužadinimą dėl jonų transportavimo pokyčių fotoreceptoriuje. Vėliau rodopsinas atkuriamas (regeneruojamas) dėl 11-ųjų sintezės. cis-tinklainė ir opsinas arba naujų išorinio tinklainės sluoksnio diskų sintezės procese.
Šiuo metu padaryta tam tikra pažanga siekiant suprasti, kas vyksta paskutiniame sužadinimo kaskados etape - ant išorinės strypų membranos. Ląstelės citoplazminė membrana yra selektyviai pralaidi elektriškai įkrautiems jonams (Na +, Ca 2+), dėl to susidaro elektrinio potencialo skirtumas tarp ląstelės membranos vidinės ir išorinės pusės. Ramybės būsenoje ląstelės membranos viduje yra apie 40 mV neigiamas krūvis išorės atžvilgiu. Aštuntajame dešimtmetyje mokslininkai parodė, kad apšvietus ląstelę šviesa, potencialų skirtumas strypo membranoje didėja. Šis padidėjimas priklauso nuo stimulo intensyvumo ir foninio apšvietimo; Didžiausias potencialų skirtumas šiuo atveju yra 80 mV.
Potencialų skirtumo padidėjimas - hiperpoliarizacija atsiranda dėl sumažėjusio membranos pralaidumo natrio katijonams Na +, kurie turi teigiamą krūvį. Nustačius hiperpoliarizacijos pobūdį, buvo nustatyta, kad dėl vieno fotono absorbcijos strypo plazminėje membranoje užsidaro šimtai natrio kanalų, užkertant kelią milijonų Na + jonų patekimui į ląstelę. Hiperpoliarizacija, atsiradusi veikiant šviesai, išilgai išorinės lazdelės membranos plinta į kitą ląstelės galą iki sinapsinio galo, kur atsiranda nervinis impulsas ir perduodamas į smegenis.
Šie esminiai tyrimai suteikė įžvalgos apie tai, kas vyksta fotocheminės vizualinio šviesos suvokimo kaskados pradžioje ir pabaigoje, tačiau liko neišspręstas klausimas, kas vyksta viduryje? Kaip tinklainės molekulės izomerizacija lazdelės disko membranoje sukelia natrio kanalų uždarymą išorinėje ląstelės membranoje? Kaip žinoma, strypuose plazminė membrana nesiliečia su disko membrana. Tai reiškia, kad signalo perdavimas iš diskų į išorinę membraną turi būti atliekamas naudojant intracelulinį sužadinimo signalo tarpininką. Kadangi vienas fotonas gali uždaryti šimtus natrio kanalų, kiekvieną fotonų sugerties įvykį turi lydėti daugybė pasiuntinių molekulių.
1973 metais buvo pasiūlyta, kad tamsoje kalcio jonai Ca + kaupiasi diskuose, o apšviesti jie išsiskiria ir, difuzijos būdu pasiekę plazmos membraną, uždaro natrio kanalus. Ši patraukli hipotezė sukėlė didelį susidomėjimą ir paskatino daugybę eksperimentų. Tačiau vėlesni eksperimentai parodė, kad nors kalcio jonai Ca + vaidina svarbų vaidmenį regėjimui, jie nėra sužadinimo siųstuvas. Tarpininko vaidmenį, kaip paaiškėjo, atlieka 3", 5"-ciklinis guanozino monofosfatas (cGMP) (7 pav.).

CGMP gebėjimą veikti kaip tarpininku lemia jo cheminė struktūra. cGMP yra guanilo nukleotidų klasės nukleotidas, randamas RNR. Kaip ir kiti nukleotidai, jis susideda iš dviejų komponentų: azoto bazės, guanino, ir penkių anglies cukraus liekanos ribozės, kurios anglies atomai 3" ir 5" padėtyse yra sujungti per fosfato grupę. Fosfodiesterio jungtis uždaro cGMP molekulę į žiedą. Kai šis žiedas yra nepažeistas, cGMP gali išlaikyti atvirus membranos natrio kanalus, o kai fosfodiesterio jungtis suskaidoma fermentu fosfodiesteraze, natrio kanalai spontaniškai užsidaro, todėl pasikeičia membranos elektrinės savybės ir gali atsirasti nervinis impulsas (8 pav.).

Tarp rodopsino sužadinimo ir fermentinio cGMP skilimo yra keli tarpiniai etapai. Kai molekulė yra 11- cis-tinklainė sugeria fotoną ir suaktyvinamas opsinas, o rodopsinas savo ruožtu aktyvuoja fermentą, vadinamą transducinu. Aktyvintos rodopsino formos sąveika su G baltymo transducinu yra pagrindinis biocheminis regėjimo proceso etapas. Transducinas yra pagrindinis tarpinis sužadinimo kaskados produktas. Šis receptoriaus G baltymas aktyvuoja specifinę fosfodiesterazę, kuri atidaro cGMP žiedą, prie jo prijungdama vandens molekulę, hidrolizuoja cGMP. Nors šio proceso schemą apibūdinti nesunku, jo fiziologiniam vaidmeniui išsiaiškinti ir suprasti prireikė daugybės įvairių eksperimentų.
Vėliau buvo nustatyta, kad cGMP koncentracija išoriniuose strypų segmentuose šviesoje mažėja. Vėlesni eksperimentai parodė, kad šis sumažėjimas yra cGMP hidrolizės, atliekamos šiam nukleotidui būdingos fosfodiesterazės, pasekmė. Tuo metu kalcio hipotezė vis dar buvo labai populiari, tačiau nebeliko abejonių, kad cGMP turėjo reikšmingos tiesioginės įtakos sužadinimo reakcijai.
1978 metais vykusioje konferencijoje P. Liebmanas iš Pensilvanijos universiteto pranešė, kad lazdelės išorinių segmentų suspensijoje vienas fotonas gali inicijuoti šimtų fosfodiesterazės molekulių aktyvavimą per sekundę. Ankstesniame darbe buvo pastebėtas daug mažesnis pagerėjimas, kai yra kitas nukleotidas, adenozino trifosfatas (ATP), nei esant guanozino trifosfatui (GTP).
Guanozino trifosfatas (GTP) turi tokią pačią struktūrą kaip ir neciklinė GMP forma, tačiau GMP 5" anglies atomas yra susietas ne su viena fosfatų grupe, o su trijų fosfatų grandine, sujungta viena su kita fosfodiesterio ryšiais. šiuose ryšiuose sukaupta energija naudojama daugeliui ląstelių funkcijų.Pavyzdžiui, iš GTP pašalinus vieną fosfatų grupę (susidarant guanozindifosfatui, BVP), išsiskiria nemažas energijos kiekis.Tokiu būdu ląstelė gauna energijos, kuri leidžia Taip pat svarbu tai, kad šis procesas vyksta aktyvavus fosfodiesterazę, kur GTP yra būtinas kofaktorius.
1994 m. buvo galima suleisti cGMP į nepažeistos lazdelės išorinį segmentą, o rezultatai buvo įspūdingi. Kai tik ciklinis guanozino monofosfatas pateko į ląstelę, potencialų skirtumas plazmos membranoje greitai sumažėjo, o vėlavimas tarp šviesos impulso taikymo ir membranos hiperpoliarizacijos smarkiai padidėjo. Taip yra todėl, kad cGMP atveria natrio kanalus ir jie lieka atviri tol, kol cGMP šviesa aktyvuota fosfodiesterazė suskaido į GMP. Ši hipotezė atrodė labai patraukli, tačiau tiesioginių jos įrodymų nebuvo.
Šviesos signalo perdavimo mechanizme didelę reikšmę turi tai, kad fosfodiesterazei aktyvuoti reikalingas GTP. Tai rodo, kad tam tikras GTP surišantis baltymas gali būti svarbus aktyvinimo tarpinis produktas. Reikėjo atidžiai ištirti, kas atsitinka su GTP strypuose. Pirmųjų eksperimentų tikslas buvo nustatyti GTP ir jo darinių surišimą išoriniuose strypų segmentuose. Radioaktyviai pažymėtas anglies izotopas 14 C GTP buvo inkubuojamas su lazdelėmis ir jų išorinių segmentų fragmentais. Po kelių valandų vaistas buvo nuplaunamas ant filtro, kuris išlaiko membranos fragmentus ir dideles molekules, tokias kaip baltymai, ir leidžia mažoms molekulėms, įskaitant GTP ir metaboliškai susijusius junginius, praeiti. Paaiškėjo, kad didelė radioaktyvumo dalis lieka susijusi su membranos frakcija. Vėliau paaiškėjo, kad membranoje lieka ne GTP, o BVP.
Šie eksperimentai parodė, kad lazdelių membranose yra baltymas, galintis surišti GTP ir pašalinti iš jo vieną fosfatų grupę, kad susidarytų BVP. Vis labiau atrodė, kad toks baltymas yra pagrindinis tarpinis produktas ir kad GTP pavertimas BVP gali paskatinti aktyvavimo procesą.
Vienas iš stulbinančių faktų buvo tai, kad lazdelių membranos ne tik suriša guanilo nukleotidus, bet, kai apšviečiamos, iš jų išsiskiria BVP, o tai procesas, kurį žymiai sustiprina GTP buvimas tirpale. Suformuota hipotezė šiems reiškiniams paaiškinti. Matyt, kai kurie aktyvinimo proceso žingsniai apima GTP keitimą į BVP membranoje. Štai kodėl BVP išleidimas yra toks stiprus ir didėja, kai pridedamas GTP: GTP turi būti pakeistas BVP. GTP vėliau virsta BVP.
Nustatyta, kad GTP keitimas į BVP yra susijęs su centriniu aktyvinimo proceso įvykiu. Ištirtas šviesos poveikis BVP absorbcijai lazdelių membranose ir nustatyta, kad vienos rodopsino molekulės fotosužadinimas lemia apie 500 GTP molekulių surišimą. Šio patobulinimo atradimas buvo svarbus žingsnis siekiant paaiškinti patobulinimą, būdingą sužadinimo kaskadai.
Šis esminis rezultatas leido padaryti svarbią išvadą, kad sužadinimo kaskadoje yra baltymų tarpinis produktas, kuris egzistuoja dviejose būsenose. Vienoje valstybėje jis suriša BVP, kitoje – GTP. BVP keitimą į GTP, kuris yra baltymų aktyvavimo signalas, inicijuoja rodopsino molekulė ir savo ruožtu aktyvuoja specifinę fosfodiesterazę. Fosfodiesterazė skaldo ciklinį GMP, kuris uždaro natrio kanalus plazmos membranoje. Šis baltymas netrukus buvo išskirtas. Jis vadinamas transducinu, nes jis tarpininkauja transdukcijai – šviesos pavertimui elektriniu signalu. Nustatyta, kad transducinas susideda iš trijų baltymų subvienetų - alfa (α), beta (β) ir gama (γ).
Signalas perduodamas iš aktyvuoto rodopsino į transduciną ir iš jo GTP formos į fosfodiesterazę. Jei šis vaizdas teisingas, galima tikėtis, kad, pirma, transducinas gali būti paverstas GTP forma, kai nėra fosfodiesterazės, ir, antra, kad fosfodiesterazę gali suaktyvinti šviesos sužadintas rodopsinas. Norint patikrinti šią prielaidą, buvo naudojama sintetinė membraninė sistema, kurioje nėra fosfodiesterazės. Ant dirbtinės membranos buvo užteptas išgrynintas BVP formos transducinas, o po to pridėtas aktyvuotas rodopsinas. Šių eksperimentų metu buvo nustatyta, kad kiekviena rodopsino molekulė katalizuoja 71 GTP analoginės molekulės įsisavinimą į membraną. Tai reiškia, kad aktyvindama transduciną, kiekviena rodopsino molekulė katalizuoja BVP keitimą į GTP daugelyje transducino molekulių. Taigi buvo galima atrasti sustiprinantį rodopsino poveikį, kurio pasireiškimui buvo išskirta išgryninta aktyvi transducino forma - jo komplekso su GTP pavidalu. Čia tyrinėtojų laukė staigmena. Neaktyvioje BVP formoje transducino molekulė yra nepažeista – visi trys jos subvienetai išsidėstę kartu. Paaiškėjo, kad pereinant prie GTP formos, transducinas disocijuoja: α subvienetas atsiskiria nuo baltymo β ir γ subvienetų, o GTP prisijungia prie laisvo α subvieneto.
Reikėjo išsiaiškinti, kuris transducino subvienetas - α- (su prijungtu GTP) arba β-, γ-subvienetas aktyvina fosfodiesterazę. Nustatyta, kad fosfodiesterazę aktyvuoja α subvienetas komplekse su GTP; kartu likę β ir γ subvienetai neturi įtakos fermento funkcionavimui. Be to, α-subvienetas sukėlė transducino aktyvavimą net ir be rodopsino; tai paaiškino prielaidą, kad transducinas gali aktyvuoti fosfodiesterazę be rodopsino.
Dabar buvo išsamiai ištirtas specifinės fosfodiesterazės aktyvinimo transducinu mechanizmas. Tamsoje fosfodiesterazė turi mažai aktyvumo, nes ji yra inaktyvuota. Pridėjus nedidelį kiekį tripsino, baltymus skaidančio fermento, suaktyvinama fosfodiesterazė. Fosfodiesterazės molekulė susideda iš trijų polipeptidinių grandinių; kaip ir transducinas, jie žymimi α- , β- ir γ- subvienetai . T ripsinas naikina γ - subvienetas, bet ne α- ir β -subvienetas. Taigi paaiškėjo, kad γ-subvienetas veikia kaip fosfodiesterazės inhibitorius.
Vėliau pavyko išskirti γ-subvienetą gryna forma, įtraukti į aktyvų α, β-subvienetų kompleksą ir buvo atrasta, kad γ-subvienetas slopina transducino katalizinį aktyvumą daugiau nei 99%. Be to, sunaikinimo greitis γ - Tripsino subvienetai gerai atitinka fosfodiesterazės aktyvacijos greitį sužadinimo kaskadoje. Transducinas GTP formoje gali prisijungti prie γ - fosfodiesterazės subvienetas, sudarantis kompleksą.
Visi šie duomenys pateikiami toliau pateiktame paveikslėlyje. Po šviesos poveikio transducino α-subvienetas su prijungtu GTP prisijungia prie fosfodiesterazės, o jį slopinantis γ-subvienetas išsiskiria. Dėl to suaktyvinamas transducinas ir pasireiškia katalizinis fosfodiesterazės aktyvumas. Šis aktyvumas yra puikus: kiekviena aktyvuota fermento molekulė per 1 sekundę gali hidrolizuoti 4200 ciklinio guanozino monofosfato molekulių. Taigi paaiškėjo dauguma regėjimo ciklo biocheminių reakcijų (9 pav.). Pradinis sužadinimo kaskados etapas yra fotono sugertis rodopsinu. Tada aktyvuotas rodopsinas sąveikauja su transducinu, dėl kurio BVP keičiasi į GTP, kuris atsiranda transducino α-subvienete. Dėl to α subvienetas yra atskirtas nuo likusio fermento, aktyvuojant fosfodiesterazę. Pastarasis suskaido daugybę cGMP molekulių . Šis procesas trunka tik apie milisekundę. Po kurio laiko transdukino α-subvieneto „įtaisytas laikmatis“ suskaido GTP, kad susidarytų BVP, o α-subvienetas vėl sujungiamas su β- ir γ-subvienetais. . Taip pat atkuriama fosfodiesterazė. Rodopsinas inaktyvuojamas, o po to pasikeičia į aktyvavimui paruoštą formą.

Veikiant vienai rodopsino molekulei, susidaro keli šimtai aktyvių α kompleksų - GTP transducino subvienetas, kuris yra pirmasis amplifikacijos žingsnis. Transducino α-subvienetas, turintis GTP, aktyvina fosfodiesterazę. Šiame etape nėra stiprinimo; Kiekviena transducino α-subvieneto molekulė jungiasi ir aktyvuoja vieną fosfodiesterazės molekulę. Kitą amplifikacijos etapą užtikrina transducino-fosfodiesterazės pora, veikianti kaip viena. Transducino α-subvienetas lieka susietas su fosfodiesteraze, kol jis suskaido 3 "-5" ryšį cikliniame guanozino monofosfate. Kiekviena aktyvuota fermento molekulė gali paversti kelis tūkstančius GMP molekulių. Šis rodopsino suteikiamas stiprinimas yra nepaprasto konversijos efektyvumo pagrindas, kai vienas fotonas sukelia intensyvų nervinį impulsą.
Tačiau kūnas sugeba daug kartų suvokti šviesą, o tai reiškia, kad šis ciklas turi išsijungti. Pasirodo, transducinas atlieka pagrindinį vaidmenį ne tik aktyvuojant, bet ir deaktyvuojant. Jo α-subvienetas turi įmontuotą "laikmačio" mechanizmą, kuris pertraukia aktyvuotą būseną, paversdamas susietą GTP į BVP. Šio „laikmačio“ veikimo mechanizmas nėra visiškai aiškus. Yra žinoma, kad GTP hidrolizė su BVP susidarymu dezaktyvavimo fazėje vaidina svarbų vaidmenį įgyvendinant visą ciklą. Reakcijos, vedančios į aktyvavimą, yra energetiškai palankios. Priešingai, kai kurios deaktyvavimo reakcijos yra nepalankios; Nekonvertavus GTP į BVP, sistema negali būti iš naujo nustatyta naujam aktyvinimui.
Kai GTP suskaidomas, kad susidarytų BVP, transducino α subvienetas išskiria slopinamąjį fosfodiesterazės γ subvienetą. Tada γ subvienetas vėl prisijungia prie fosfodiesterazės, grąžindamas ją į ramybės būseną. Transducinas atkuria savo išankstinę aktyvavimo formą dėl subvienetų α ir β, γ susijungimo . Rodopsiną deaktyvuoja fermentas – kinazė, kuri atpažįsta specifinę jo struktūrą. Šis fermentas prideda fosfatų grupes prie kelių aminorūgščių viename opsino polipeptidinės grandinės gale. Tada rodopsinas sudaro kompleksą su baltymu arrestinu, kuris blokuoja transducino prisijungimą ir grąžina sistemą į tamsią būseną.
Vizualinės kaskados tyrimai devintojo dešimtmečio viduryje ir dešimtojo dešimtmečio pradžioje. labai rėmėsi prielaida, kad ciklinis guanozino monofosfatas atveria natrio kanalus išorinėje lazdelės membranoje ir kad dėl jo hidrolizės jie užsidaro. Tačiau apie šių procesų mechanizmus buvo žinoma mažai. Ar cGMP veikia kanalus tiesiogiai ar per tam tikrus tarpinius veiksmus? Tikslų atsakymą į šį klausimą 1985 metais gavo rusų mokslininkas E.E. Fesenko iš Biologinės fizikos instituto Maskvoje. Eksperimentuose buvo naudojama mikropipetė, į kurią buvo įtraukta nedidelė lazdelės plazminės membranos dalis. Jis tvirtai prilipo prie pipetės galiuko, o ta pusė, kuri paprastai būtų nukreipta į ląstelės vidų, pasirodė esanti išorė. Ši membranos pusė buvo plaunama įvairiais tirpalais ir nustatyta jų įtaka natrio laidumui. Rezultatai buvo visiškai nedviprasmiški: natrio kanalai atsidaro tiesiogiai su cGMP; kitos medžiagos, įskaitant kalcio jonus Ca +, jų neveikia.
Puikūs Rusijos mokslininkų eksperimentai paneigė kalcio jonų Ca + kaip sužadinimo tarpininko idėją ir nustatė paskutinę sužadinimo kaskados grandį. Taip pat paaiškėjo bendri sužadinimo grandinės kontūrai. Kaip ir tikėtasi, informacijos srautas yra iš rodopsino į transduciną, tada į fosfodiesterazę ir galiausiai į cGMP.
Nors sužadinimo kaskados būdų ir mechanizmų tyrimas padarė didelę pažangą, daugelis svarbių klausimų vis dar lieka neatsakyti. Visų pirma, neaišku, kaip reguliuojamas kaskados stiprinimo atsakas. Strypai yra daug mažiau jautrūs ryškioje šviesoje nei tamsoje. Foninis apšvietimas turi kažkaip paveikti bendrą sistemos rezultatą, ty bendrą stiprinimą, sukuriamą dviem etapais - perduodant signalą iš rodopsino į transduciną ir iš fosfodiesterazės į cGMP. Daug įrodymų rodo, kad šiame procese dalyvauja kalcio jonai, tačiau šio mechanizmo detalės nėra visiškai suprantamos. Šiuo atžvilgiu taip pat buvo svarbu nustatyti natrio kanalų struktūrą ir mechanizmus, užkertančius kelią ciklinio guanozino monofosfato išeikvojimui ląstelėje. Didelį indėlį į šio tyrimo tyrimą įnešė B. Kauppo iš Osnabriuko universiteto (Vokietija) Neurobiologijos instituto ir Liebmanno grupės: jie išskyrė cGMP valdomus kanalus ir atkūrė jų funkciją modelio membranose. Pagrindinis elementas yra guanilato ciklazė, fermentas, sintetinantis cGMP. Ląstelėje yra grįžtamojo ryšio tipo cGMP koncentracijos reguliavimas, užtikrinantis, kad po reakcijos į šviesos dirgiklį cGMP koncentracija atsistato iki pradinio lygio. Be to ląstelė turėtų galimybę dirbti tik keletą kartų ir taip išnaudotų savo gebėjimą reaguoti ilgą laiką.
Naujausių vizualinių reakcijų kaskados lazdelėse tyrimų rezultatai yra svarbūs ir kitų tipų ląstelėms. Šviesos signalo konvertavimo sistema kitose fotoreceptorių ląstelėse – kūgiuose – panaši į strypų. Yra žinoma, kad kūgiuose yra trys į rodopsiną panašūs vizualiniai pigmentai, reaguojantys į tam tikro bangos ilgio šviesą – raudona, žalia arba mėlyna. Visuose trijuose pigmentuose yra 11- cis- tinklainė. Taikant molekulinės genetikos metodus, nustatyta, kad kūgio pigmentų struktūra yra tokia pati kaip rodopsino. Transducino, fosfodiesterazės ir cGMP valdomi kanalai kūgiuose ir strypuose yra labai panašūs.
Kaskados, susijusios su cikliniu guanozino monofosfatu, reikšmė neapsiriboja regėjimu. Sužadinimo kaskados strypuose turi pastebimą panašumą su kai kurių hormonų veikimo mechanizmu. Pavyzdžiui, adrenalinas veikia aktyvuodamas fermentą, vadinamą adenilato ciklaze. Adenilato ciklazė katalizuoja ciklinio adenozino monofosfato (cAMP), kuris yra daugelio hormonų tarpląstelinis pasiuntinys, susidarymą. Buvo aptiktas ryškus šios reakcijos panašumas su sužadinimo kaskados veikimu strypuose. Lygiai taip pat, kaip sužadinimo kaskada prasideda nuo fotono absorbcijos rodopsinu, hormonų kaskada prasideda nuo hormono prisijungimo prie specifinio baltymo receptoriaus, esančio ląstelės paviršiuje. Receptoriaus-hormono kompleksas sąveikauja su vadinamuoju G baltymu, kuris primena transduciną. Tas pats surištų molekulių keitimas, kuris aktyvuoja transduciną (GTP į BVP), taip pat suaktyvina G baltymą, kai jis sąveikauja su receptoriaus-hormono kompleksu. G baltymas, kaip ir transducinas, susideda iš trijų subvienetų. Adenilato ciklazę aktyvuoja jos α-subvienetas, kuris pašalina slopinamąjį poveikį. G baltymo stimuliuojantis poveikis taip pat nutrūksta dėl integruoto „laikmačio“, kuris paverčia GTP į BVP.
Transducino ir G baltymų panašumas taikomas ne tik aktyvumui, bet ir struktūrai. Transducinas ir G baltymai priklauso tai pačiai šeimai – receptorių membraninių baltymų, perduodančių tam tikrus signalus, šeimai. Visi iki šiol nustatyti šios grupės atstovai turi beveik tą patį α-subvienetą. Be to, α subvienetas atlieka tą pačią funkciją, kaip parodyta molekuliniu lygiu. Neseniai kelios laboratorijos nustatė DNR nukleotidų sekas, koduojančias transducino α-subvienetus ir tris G baltymus. Sprendžiant iš DNR, šių keturių polipeptidinių grandinių aminorūgščių sekos yra identiškos arba beveik identiškos viena kitai maždaug pusę jų ilgio.
Lyginamoji genetinės informacijos analizė atskleidė, kad transducino ir G-baltymų α-subvienetuose yra tiek evoliucijos metu nepakitusių, tiek stipriai besiskiriančių regionų. Kiekvienas baltymas turi tris surišimo vietas: vieną guanilo nukleotidams, vieną aktyvuotam receptoriui (rodopsinui arba hormonų receptorių kompleksui) ir vieną efektorinio baltymo fosfodiesterazei arba adenilato ciklazei. GTP ir BVP surišimo vietos, kaip būtų galima tikėtis atsižvelgiant į jų lemiamą vaidmenį sužadinimo kaskadoje, pasirodė esančios labiausiai išsaugotos.
Be to, paaiškėjo, kad šių baltymų GTP surišančios sritys primena vieną funkciškai visiškai kito baltymo regioną; vadinamasis pailgėjimo koeficientas Tu. Šis baltymas vaidina svarbų vaidmenį baltymų sintezėje: sudaro kompleksą su GTP ir aminoacil-tRNR molekulėmis, o vėliau jungiasi prie ribosomos, t.y. užtikrina pailgėjimo procesą – aminorūgščių patekimą į susintetinto augimo vietą. polipeptidinė grandinė. Įvykių ciklas, vykstantis su Tu baltymu jo veikimo metu, yra panašus į transducino ciklą. Ciklas prasideda nuo GTP skilimo. Tu molekulėje yra GTP surišimo vieta, o aminorūgščių seka ji labai panaši į guanilo nukleotidų surišimo vietas transducine ir įvairiuose G baltymuose.
Baltymų sintezė yra pagrindinis ląstelių metabolizmo aspektas ir tikėtina, kad pailgėjimo faktorius Tu, kuris dalyvauja šiame pagrindiniame procese, išsivystė anksčiau nei G baltymai ar su jais susijęs transducinas. Šis įdomus baltymas gali būti tiek transducino, tiek G baltymų protėvis. Kontroliuojamas baltymų išsiskyrimas ir surišimas, susijęs su GTP keitimu į BVP, susidarė ankstyvoje evoliucijos stadijoje, o pailgėjimo faktorius Tu gali būti vienas iš pirmųjų tokio ciklo evoliucinių variantų.
Vienas iš nuostabiausių evoliucijos dalykų yra tai, kad tam tikrai funkcijai atsiradęs mechanizmas vėliau gali būti modifikuotas ir naudojamas visiškai kitoms funkcijoms. Būtent taip atsitiko su Tu veikimo mechanizmu. Evoliucijos metu susiformavęs baltymų sintezei vykdyti, jis išliko milijardus metų, o vėliau pateko į hormoninių ir sensorinių signalų perdavimo sistemą. Per pastaruosius kelerius metus viena iš jo funkcijų – transducino ciklas – buvo labai išsamiai ištirta. Šių tyrimų rezultatai turi didelę mokslinę reikšmę, nes molekuliniu lygmeniu buvo galima suprasti vieną nuostabiausių jutimo mechanizmų – šviesos perdavimo ir vizualinės stimuliacijos mechanizmą.
Galbūt netrukus bus atskleistos naujos idėjos apie spalvinį matymą. Vis dar neaišku, ar žalia spalva, kurią matome, yra vidurinis efektas tarp geltonos ir mėlynos, ar kai kuriais atvejais ji atitinka bangos ilgius, atitinkančius žalią spektro spalvą.
Mūsų smegenys gali užregistruoti žalią spalvą kaip spektrometras, ty esant tam tikram elektromagnetinių bangų ilgiui. Jis taip pat gali registruoti žalią kaip geltonos ir mėlynos spalvos mišinį. Spalvų suvokimas vizualiniu analizatoriumi negali būti nustatytas kaip spektrometras.
Kaip elektromagnetinių bangų, atitinkančių žalią ir raudoną, maišymosi pavyzdys pateikta geltona spalva. Manoma, kad vizualinio veiksmo metu veikia mėlynos-geltonos ir žalios-raudonos spalvų poros. Vaizdinis analizatorius turi galimybę analizuoti tam tikrus optinio spektro diapazonus, pavyzdžiui, spalvas. Sumaišius žalią ir raudoną, vidurinė spalva negaunama. Smegenys jį suvokia kaip geltoną. Kai skleidžiamos elektromagnetinės bangos, atitinkančios žalią ir raudoną, smegenys suvokia „vidurinį tirpalą“ - geltoną.

Lygiai taip pat mėlyna ir geltona suvokiama kaip žalia. Tai reiškia, kad tarp porų mėlyna-geltona ir žalia-raudona atsiranda spektrinis spalvų maišymasis. Tai taip pat taikoma situacijai, kai vizualinis analizatorius „priima sprendimą“ dėl spalvų, kurioms jis yra jautresnis. Taip pat žalia ir mėlyna spalvos suvokiamos kaip žalsvai mėlynos spalvos. Pavyzdžiui, vizualinis analizatorius oranžinę spalvą visada suvokia kaip oranžinę, nes nuo jos atsispindi elektromagnetinės bangos, atitinkančios geltoną ir raudoną spalvą. Mažiausias regėjimo jautrumas yra violetinei, mėlynai ir raudonai. Be to, elektromagnetinių bangų mišinys, atitinkantis mėlyną ir raudoną spalvas, yra suvokiamas kaip violetinis. Kai sumaišomos elektromagnetinės bangos, atitinkančios daugiau spalvų, smegenys jas suvokia ne kaip atskiras spalvas ar kaip „vidutinį“ sprendimą, o kaip baltas. Šie duomenys rodo, kad spalvos samprata nėra vienareikšmiškai nulemta bangos ilgio. Analizę atlieka „biokompiuteris“ - smegenys, o spalvos idėja iš esmės yra mūsų sąmonės produktas.
IŠVADA
Medicinos mokslų tyrimų centre (Bulgarija) pastaruosius 10 metų vykdomi rodopsino ir kitų susijusių tinklainės turinčių chromoforinių baltymų (jodopsino, bakteriorodopsino) struktūriniai tyrimai, taip pat su jo funkcionavimu susijusių akių patologijų nustatymas. ir tarp problemų, kurias reikia skubiai išspręsti, galima išskirti:
Kokios struktūrinės transformacijos lydi rodopsino aktyvavimą ir suteikia jam galimybę sąveikauti su receptorių G baltymais (transducinu, proteinkinazėmis ir arrestinu)?
Kokios yra aktyvuoto rodopsino ir transducino kompleksų erdvinės struktūros?
Koks yra ląstelių „brendimo“ ir rodopsino skilimo mechanizmas?
Tolesni rodopsino tyrimai turi ne tik fundamentalią mokslinę, bet ir taikomąją reikšmę, gali būti naudojami biocheminiams regos sutrikimams gydyti ar jų profilaktikai. Rodopsinas yra labiausiai ištirtas GPCR receptorių šeimos baltymas, o aukščiau pateikti rezultatai gali būti naudojami tiriant kitų šios šeimos transmembraninių baltymų, pavyzdžiui, bakteriorodopsino, struktūrą ir funkcines savybes.
1. D. Hubelis. Akys, smegenys, regėjimas/ red. A. L. Byzova., Mir, Maskva (1990), 172 p.
2. M. J. Hoganas, J. A. Alvarado, J. E. Weddellas. Žmogaus akies histologija, Saunders, Filadelfija (1971), 115 p.
3. J. Nathans, D. Thomas, D. S. Hogness. “ Žmogaus spalvinio matymo molekulinė genetika: genai, koduojantys mėlyną, žalią ir raudoną pigmentus“, Mokslas, 232(47), 193–202 (1986).
4. R. Henderson, J. M. Baldwin, T. A. Ceska, F. Zemlin, E. Beckmann, K. H. Downing. „Bakteriorodopsino struktūros modelis, pagrįstas didelės skiriamosios gebos elektronų kriomikroskopija“, J. Mol. Biol., 212 , 899–29 (1991).
5. K. Palczewski, T. Kumasaka, T. Hori, C. A. Behnke, H. Motoshima, B. A. Fox, I. Le Trong, D. C. Teller, T. Okada, R.E. Stenkamp, M. Yamamoto, M. Miyano, „Rhodopsino kristalinė struktūra: su G baltymu susietas receptorius“, Mokslas, 289 , 739–745 (2000).
6. Ju. A. Ovčinikovas, N. G. Abdulajevas, M. Ju. Feigina, I. D. Artamonovas, A. S. Bogačukas. „Vizualinis rodopsinas: visa aminorūgščių seka ir topologija membranoje“, Bioorganinė chemija, 10 , 1331–1340 19830.
7. P.A. Hargrave'as, J.H. McDowell, D.R. Curtis, J. K. Wang, E. Juszczak, S. L. Fong, J. K. Rao, P. Argos, „Galvių rodopsino struktūra“, Biofizė. Struktūra. Mech., 9 , 235–244 (1983).
8. G. F. Schertleris, P. A. Hargrave'as, „Dviejų kristalų formų varlės rodopsino projekcijos struktūra“, Proc. Natl. Akad. Sci. U. S. A., 9 2, 11578–11582 (1995).
9. V. M. Lipkinas. „Vizualinė sistema. Vaizdinio signalo perdavimo ir stiprinimo tinklainėje mechanizmai. Soroso edukacinis žurnalas, 9 , 2–8 (2001).
10. Y. Shichida, H. Imai. „Vizualinis pigmentas: su G baltymu sujungtas šviesos signalų receptorius“, Ląstelė. Mol. Gyvenimas Sci., 54 , 1299–1315 (1998).
11. A. B. Rubinas. Bakteriorodopsino ir rodopsino fototransformacijos, Biofizika, t. 2., Maskva, Nauka (2004), 87 p.
12. Y. Liang, D. Fotiadis, T. Maeda, A. Maeda, A. Modzelewska, S. Filipek, D. A. Saperstein, A. Engel, K. Palczewski. „Rodopsino signalizacija ir organizavimas heterozigotinėse rodopsino išjungimo pelėse“, J. Biol. Chem., 279 , 48189–48196 (2004).
13. J. M. Baldwin, G. F. Schertler, V. M. Unger. „α anglies šablonas, skirtas su G baltymu susietų receptorių rodopsino šeimos transmembraninėms spiracijoms“, J. Mol. Biol., 272 , 144–164 (1997).
14. J. Fitzgibbon, B. Appukuttan, S. Gayther, D. Wells, J. Delhanty, D. M. Hunt. „Žmogaus mėlynojo kūgio pigmento geno lokalizavimas į 7q31.3-32 chromosomos juostą“, Žmogaus genetika, 93 (1), 79–80 (1994).
15. K. Palczewski „G-baltymų prijungtas receptorius rodopsinas“, Annu. Rev. Biochem., 7 5, 743–767 (2006).
16. P. S. Park, S. Filipek, J. W. Wells, K. Palczewski. „Su G baltymu susietų receptorių oligomerizacija: praeitis, dabartis ir ateitis“, Biochemija, 43 , 15643–15656 (2004).
17. I. Ignatovas, M. Marinovas. Spalvų Kirliano spektrinė analizė. Spalvų stebėjimas naudojant vizualinį analizatorių, EUROMEDICA, Hanoveris, (2008), 32 p.
18. O.V. Mosinas, I. I. Ignatovas. „Natūralus fotokonvertuojantis nanomedžiagos bakteriorodopsinas iš halofilinės bakterijos Halobacterium halobium“, Nanomedžiagos ir nanostruktūros, 2 , 47-58 (2012).

Kokie būtų Naujieji metai be šampano, mandarinų, Olivier, aspicų ir visų mėgstamo „Silkės po kailiu“. Su paskutiniu...

Paruoškime sausainiams reikalingus ingredientus. Pirmas dalykas, kurį reikia padaryti, yra užvirti vandenį. Mums reikia...

Ar galima registruoti darbuotoją į finansų direktoriaus – vyriausiojo buhalterio pareigas? Vyriausioji buhalterė tvirtina, kad taip, bet...

Smulkaus verslo vadovas gali lengvai valdyti biudžetą savarankiškai. PATIKRINTA! Jei planuojate biudžetą...

Naujų projektų kūrimas apima preliminarų ekonominį jų pagrįstumo pagrindimą, vėlesnius...
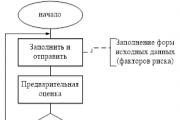
Ataskaitas rengia RM, dėl jų susitaria (patvirtina) Rizikos komitetas prie valdybos ir perduoda...

Didelės, labai didelės pievos pakraštyje, ant ilgo smaragdinio žolės stiebo gyveno mažytė boružėlė. Dievo mažutė...

Šiais laikais gana įprasta, kad žmonės atsigręžia į žvaigždes. Horoskopo pagalba žmogus gali sužinoti...

Verslo ar draugiško. Jei persekiojote nepažįstamą žmogų, tai reiškia jūsų pasitikėjimo moteriškąja lytimi lygį...

pagal Freudo svajonių knygą Jei svajojote, kaip žvejojote, tai reiškia, kad realiame gyvenime vargu ar galite išsijungti...

Naujųjų metų viesulas suks mus taip greitai ir greitai, kad laikas gerai apgalvoti Naujųjų metų...

Straipsnyje pateikiami keli skaniausi vinaigretės gaminimo receptai. Vinaigrette yra skanios salotos...

2015 m. liepos 4 d., 20:33 Paskutiniu metu mūsų Nastena visiškai atsisako saldumynų (ir ačiū Dievui), bet...

Kvapus, labai skanus šokoladinis pyragas, neįmanoma nustoti valgyti! Tortą Negro’s Kiss labai lengva paruošti,...

Paruoškime sausainiams reikalingus ingredientus. Pirmas dalykas, kurį reikia padaryti, yra užvirti vandenį. Mes...

Ar galima registruoti darbuotoją į finansų direktoriaus – vyriausiojo buhalterio pareigas? Vyriausioji buhalterė tvirtina...