O que é possível e o que não é possível durante o Jejum da Natividade?
Em 2018, o Jejum da Natividade começará em 28 de novembro. Durante este período, os crentes ortodoxos se preparam para celebrar o Natal...
Grupos C e G. A proteína G é semelhante à proteína A, mas difere em especificidade. A proteína G tem peso molecular de 58 kDa (no caso da proteína C40) ou 65 kDa (no caso da proteína G148). A proteína G liga-se à região α dos anticorpos e, portanto, é amplamente utilizada para a purificação de imunoglobulinas. As moléculas de proteína G também se ligam à albumina.
Existem outras proteínas bacterianas que se ligam às imunoglobulinas - proteína A, proteína A/G e proteína L. Estas proteínas são utilizadas para purificação, imobilização e isolamento de imunoglobulinas. Estas proteínas de ligação a anticorpos têm diferentes perfis de ligação a imunoglobulinas.
Fundação Wikimedia. 2010.
- (proteína A inglesa) é uma proteína com peso molecular de 40-60 kDa, isolada da superfície da parede celular de Staphylococcus aureus (Staphylococcus aureus). A proteína A é usada em pesquisas bioquímicas, pois se liga bem a muitos... ... Wikipedia
Fique boquiaberto com os esquilos árabes. Dicionário de sinônimos russos e expressões semelhantes. sob. Ed. N. Abramova, M.: Dicionários Russos, 1999. proteína globulina, histona, proteinóide, proteína, corpo protéico, protamina, proteína Dicionário de sinônimos russos ... Dicionário de sinônimo
Dicionário Explicativo de Ushakov
1. PROTEÍNA1, esquilo, masculino. (biol. química). O componente mais importante do corpo de animais e plantas; o mesmo que albumina e proteína. 2. PROTEÍNA2, esquilo, masculino. 1. Concha convexa e opaca do olho. || apenas plural Olhos (simples). Projete os brancos. Proteínas... ... Dicionário Explicativo de Ushakov
1. PROTEÍNA1, esquilo, masculino. (biol. química). O componente mais importante do corpo de animais e plantas; o mesmo que albumina e proteína. 2. PROTEÍNA2, esquilo, masculino. 1. Concha convexa e opaca do olho. || apenas plural Olhos (simples). Projete os brancos. Proteínas... ... Dicionário Explicativo de Ushakov
proteína C- Proteína, serina protease, sintetizada pelas células do fígado; anticoagulante, é um inibidor dos fatores de coagulação sanguínea Va e VIIIa; a frequência de heterozigotos para deficiência de B.C em populações humanas chega a 1/200; esta anomalia está associada ao aumento... ... Guia do Tradutor Técnico
PROTEÍNA, um COMPOSTO orgânico contendo muitos AMINOÁCIDOS conectados por ligações peptídicas covalentes. As moléculas de proteína consistem em cadeias polipeptídicas. Existem cerca de 20 aminoácidos diferentes nas CÉLULAS vivas. Devido ao fato de que em cada... ... Dicionário enciclopédico científico e técnico
PROTEÍNA, também, marido. Uma substância orgânica de alto peso molecular que garante as funções vitais dos organismos animais e vegetais. | adj. proteína, ah, ah. Alimentação proteica (alto teor de proteína). II. PROTEÍNA, também, marido. 1. A parte transparente do ovo... Dicionário Explicativo de Ozhegov
A proteína de ligação a ácidos graxos (FABP) é uma família de transportadores de ácidos graxos e outras substâncias lipofílicas, como eicosanóides e retinóides. Acredita-se que estes... ... Wikipedia
1. PROTEÍNA, lka; m. 1. O líquido transparente que envolve a gema de um ovo de pássaro. / Sobre esta parte de um ovo de galinha como alimento. Beba cru b. Brancos batidos. ◁ Proteína, ah, ah. B. creme (de clara de ovo). 2. PROTEÍNA veja 1. Proteínas. 3. PROTEÍNA veja 2. Proteínas. 4.… … dicionário enciclopédico
(Inglês) Proteínas de ligação a nucleotídeos de guanina, As proteínas de ligação ao nucleotídeo guanil são uma família de proteínas envolvidas na sinalização de células eucarióticas. As proteínas G desempenham o papel de uma espécie de interruptor: podem passar de um estado inativo para um estado ativo e vice-versa, ligando ou desligando a transmissão de um determinado sinal dentro da célula. Essas proteínas receberam esse nome por sua capacidade de se ligar a nucleotídeos guanil. Nucleotídeo guanina): em complexo com guanosina difosfato (GDP) são inativos e em complexo com guanosina trifosfato (GTP) são ativos.
O termo "proteínas G" é mais frequentemente usado para se referir a proteínas heterotriméricas (grandes) de ligação a GTP que consistem em três subunidades α, β e γ; Existe outra classe de proteínas de ligação a GTP - monômeros, às vezes chamados de pequenas proteínas G (supercasa das pequenas GTPases Ras), elas são homólogas às subunidades α das grandes.
As proteínas G heterotriméricas estão envolvidas na transmissão de sinais dos receptores acoplados à proteína G. Receptores acoplados à proteína G (GPCR)- a maior classe de receptores celulares (por exemplo, em Caenorhabditis elegans seus genes ocupam 5% de todo o genoma). Nos vertebrados, eles são responsáveis pela percepção celular de uma série de hormônios e outras moléculas sinalizadoras, bem como pelos sentidos químicos (olfato e paladar) e fotorrecepção (visão). É significativo que aproximadamente metade dos medicamentos farmacêuticos conhecidos atuem através de receptores acoplados à proteína G: estes incluem medicamentos bem conhecidos, como os anti-histamínicos Claritin (loratadina) e o antidepressivo Prozac (Fluoxetina), bem como substâncias psicotrópicas, em particular heroína, cocaína e tetrahidrocanabinol (princípio ativo da maconha).
As proteínas G heterotriméricas foram descobertas por Alfred Gilman e Martin Rodbell, pelas quais receberam o Prêmio Nobel de Fisiologia ou Medicina em 1994.
As proteínas G heterotriméricas consistem em três subunidades: α, β e γ. A subunidade α contém um domínio de ligação e hidrólise de GTP que é idêntico em toda a supercasa da GTPase. A subunidade β contém 7 estruturas β organizadas como pás de hélice. A subunidade γ interage intimamente com a subunidade β; juntas elas formam uma única estrutura funcional, que pode se dissociar apenas no caso de hidrólise de proteínas. Toda a proteína G está ancorada na membrana por dois lipídios, um dos quais está covalentemente ligado ao terminal N da subunidade α, o outro ao terminal C da subunidade γ.
Receptores acoplados à proteína G Receptores acoplados à proteína G, GPCR)- a maior família de receptores celulares em eucariotos, proporcionando a percepção de hormônios, neurotransmissores, reguladores locais, além de fornecer visão, olfato e paladar em vertebrados. Cerca de 700 genes GPCR são encontrados no genoma humano, e no camundongo existem mais de 1.000 desses receptores apenas para o sentido do olfato.
As moléculas sinalizadoras que atuam como ligantes para receptores acoplados à proteína G podem ser de natureza química muito diferente: proteínas, pequenos peptídeos, lipídios, derivados de aminoácidos e semelhantes. Além disso, alguns membros desta classe de receptores, em particular a rodopsina, podem perceber fótons de luz. Às vezes, existem vários GPCRs diferentes para uma única molécula de sinalização, são expressos em diferentes tipos de células e desencadeiam diferentes vias de sinalização. Por exemplo, no corpo humano existem pelo menos 9 receptores diferentes para a adrenalina e pelo menos 14 para o neurotransmissor serotonina.
Todos os receptores acoplados à proteína G têm uma estrutura semelhante: consistem em uma única cadeia polipeptídica que atravessa a bicamada lipídica 7 vezes. Cada domínio transmembrana é representado por uma hélice α, que inclui 20-30 aminoácidos não polares. Esses domínios são interligados por alças de vários tamanhos localizadas em ambos os lados da membrana plasmática. Os GPCRs são predominantemente glicoproteínas, cujos resíduos de carboidratos estão localizados no lado da glicoproteína. Os domínios intracelulares destes receptores contêm locais de interação com proteínas G.
As proteínas G desempenham o papel de acoplar receptores celulares a certas moléculas efetoras, como enzimas ou canais iônicos, enquanto atuam como interruptores moleculares. No estado inativo, as proteínas G contêm GDP ligado à subunidade α.
A transmissão do sinal começa quando o ligante apropriado atua no receptor celular, como resultado o receptor é ativado e muda de conformação. O receptor ativado influencia a proteína G (que está permanentemente complexada com ele ou se associa após a ativação), fazendo com que a estrutura da subunidade α mude de tal forma que libera a molécula de GDP ligada. O lugar dessa molécula é rapidamente ocupado pelo GTP, o que leva à ativação da proteína G e a alterações em sua estrutura: a subunidade α perde afinidade pelo complexo βγ e ele se desintegra. Nesse estado ativado, tanto a subunidade α ligada ao GTP quanto o complexo βγ podem realizar a transmissão do sinal: ativar certas enzimas ou influenciar o estado dos canais iônicos. A subunidade α é uma GTPase e, assim que hidrolisa o GTP ligado ao GDP, é imediatamente inativada e a estrutura trimmer da proteína G é restaurada. Desta forma o som é desligado. A proteína G inativada pode interagir com uma molécula receptora subsequente e ligar novamente.
A eficiência da transmissão de um sinal específico através da proteína G depende da razão entre a concentração das formas ativa, ligada ao GTP, e inativa, ligada ao GDP. E esta relação, por sua vez, depende de dois estados: a constante de dissociação do PIB e a constante de taxa de hidrólise do GTP:
Onde
Essa relação é confirmada pelo excesso de GTP no meio, bem como por sua ligação rápida, quase instantânea, a uma molécula de proteína G “vazia” (ou seja, não associada a um nucleotídeo guanil). Neste caso, a eficiência da transmissão do sinal pode ser ajustada de uma das seguintes maneiras:
As proteínas G recebem informações de seus receptores associados, após o que ativam uma das vias de sinalização da célula.
O AMP cíclico (cAMP) é um segundo mensageiro comum que controla muitos processos em células eucarióticas. O AMPc é sintetizado com ATP pela grande enzima transmembrana adenilato ciclase e degradado pela AMPc fosfodiesterase. Muitas moléculas sinalizadoras influenciam a célula aumentando ou diminuindo a concentração de AMPc através da ativação ou inibição da adenilato ciclase. O cAMP desempenha sua função como segundo mensageiro ativando a proteína quinase dependente de cAMP (proteína quinase A, PKA), que por sua vez fosforila muitas proteínas na célula nos resíduos de serina e treonina, ativando-as ou desativando-as.
Existem dois tipos de proteínas G que influenciam as adenilato ciclases ativas: G s (eng. Estimulante)- estimulando, ativando-o e aumentando a concentração de AMPc e G i (eng. Inibitório)- inibitório, suprimindo a adenilato ciclase, mas também atua por ação direta nos canais iônicos. Exemplos de reações desencadeadas por um aumento dependente de G na concentração de AMPc são:
As proteínas G, que influenciam a sinalização celular dependente de AMPc, são alvos de toxinas bacterianas:
Estas duas toxinas são utilizadas em estudos biológicos para determinar qual resposta celular é mediada pela proteína Gs ou Gi.
Muitos receptores acoplados à proteína G atuam ativando a fosfolipase C-β (PLC-β). Esta enzima atua sobre o fosfolipídeo inositol: fosfatidilinositol 4,5 bifosfato (PI(4,5)P2 ou PIF 2), presente em pequenas quantidades no folheto interno da bicamada lipídica da membrana plasmática. Os receptores que ativam essa via de sinalização geralmente são acoplados ao bloco G q, que ativa a fosfolipase C de maneira semelhante ao bloco G s - adenilato ciclase. A fosfolipase ativada decompõe o fosfatidilinositol 4,5 bifosfato em inositol 1,4,5 trifosfato (IP 3) e diaciglicerol (DAG). Nesta fase, a via de sinalização se ramifica:
Exemplos de reações celulares de ativação da fosfolipase C-β dependente da proteína G são:
Muitas proteínas G atuam abrindo ou fechando canais iônicos, alterando assim as propriedades elétricas da membrana plasmática.
Por exemplo, uma diminuição na frequência e força das contrações cardíacas sob a influência da acetilcolina ocorre devido ao fato de os receptores muscarínicos de acetilcolina, após ativação, interagirem com o G i -bill, cuja subunidade α suprime a atividade do adenilato ciclase, enquanto o complexo βγ abre canais de potássio na membrana plasmática dos músculos das células cardíacas, devido ao qual sua excitabilidade diminui.
Outras proteínas G regulam indiretamente a atividade dos canais iônicos: por exemplo, a visão e os receptores olfativos atuam através das proteínas G, que afetam a síntese de nucleotídeos cíclicos, por sua vez fechando ou abrindo canais iônicos (canais iônicos controlados por nucleotídeos cíclicos). Por exemplo, todos os receptores olfativos estão associados à proteína Golf, que ativa a adenilato ciclase; O AMPc sintetizado abre os canais de sódio, o que leva à despolarização da membrana e à geração de um impulso nervoso (potencial receptor) que é transmitido aos neurônios.
Nos bastonetes da retina humana, a molécula sensível à luz é a rodopsina. A membrana plasmática dessas células contém um grande número de canais catiônicos controlados por cGMP. Na ausência de estimulação luminosa, o citoplasma dos bastonetes contém uma grande quantidade de cGMP, que mantém os canais catiônicos abertos. Como resultado, a membrana se despolariza periodicamente e ocorre a transmissão sináptica de impulsos aos neurônios. Após ativação pela luz, a rodopsina muda de conformação e interage com a proteína G transducina (Gt). Depois disso, sua subunidade α ativa a cGMP fosfodiesterase, que decompõe o cGMP, como resultado do fechamento dos canais catiônicos e da interrupção da transmissão sináptica. É a diminuição da frequência dos impulsos provenientes das células sensíveis à luz que é percebida pelo cérebro como uma sensação de luz.
Todas as proteínas G heterotriméricas são divididas em quatro famílias principais com base na sequência de aminoácidos da subunidade α:
| Principais famílias de proteínas G heterotriméricas baseadas na sequência de aminoácidos da subunidade α | |||
|---|---|---|---|
| Família | Alguns membros | Subunidade responsável pelo efeito | Algumas funcionalidades |
| EU | Gs | α | Ativação da adenilato ciclase, abertura dos canais de cálcio |
| Golfe | α | Ativação da adenilato ciclase em neurônios olfativos | |
| II | G eu | α | Inibição da adenilato ciclase |
| βγ | Abertura dos canais de potássio | ||
| Ir | βγ | Abertura dos canais de potássio, fechamento dos canais de cálcio | |
| α e βγ | Ativação da fosfolipase C-β | ||
| G t (transducina) | α | Ativação da fosfodiesterase cGMP em fotorreceptores de vertebrados | |
| III | Gq | α | Ativação da fosfolipase C-β |
| 4 | G 12/13 | α | Ativação de GTPases monoméricas da família Rho que regulam o citoesqueleto de actina |
(receptores acoplados à proteína C, GPCRs), transmitem um sinal de mensageiros primários para alvos intracelulares usando a cascata de proteínas efetoras GPCR-^-proteína G^. Os sinais primários para esses receptores são uma grande variedade de moléculas, por exemplo, hormônios e neurotransmissores de baixo peso molecular (como adrenalina, norepinefrina, acetilcolina, serotonina, histamina), opioides, hormônios peptídicos e proteicos (adrenocorticotropina, somaostatina, vasopressina, angiotensina). , gonadotrofina, fator de crescimento epidérmico), alguns neuropeptídeos.
A mesma série inclui muitos sinais químicos percebidos pelas células sensoriais olfativas e gustativas, e luz, cujo receptor é o pigmento das células visuais ou fotorreceptoras rodopsina.
Deve-se levar em conta que o mesmo sinal primário pode iniciar a transmissão do sinal através de vários (às vezes mais de 10) GPCRs diferentes, portanto, se o número de sinais externos para GPCRs for de várias dezenas, então mais de 200 desses receptores são conhecidos.
Com toda a sua diversidade, os GPCRs são proteínas monoméricas de membrana integral, cuja cadeia polipeptídica atravessa a membrana celular sete vezes. Em todos os casos, o sítio receptor responsável pela interação com o sinal primário está localizado no lado externo da membrana, e a região em contato com a proteína G está no seu lado citoplasmático.
O componente a jusante da cascata de transdução de sinal GPCR é representado pela proteína G. Foram encontradas cerca de 20 proteínas G diferentes, entre elas, em primeiro lugar, devem ser mencionadas G s e C, que estimulam e inibem respectivamente a adenilato ciclase; Gq, ativando a fosfolipase C; Proteínas G das células sensoriais: fotorreceptor - G t (transducina), olfativo - G o if e gustativo - G g.
As proteínas C são heterotrímeros que consistem em três tipos de subunidades: ct, (S e y), mas em condições naturais as duas últimas subunidades funcionam como um único complexo de Ru. A característica mais importante das proteínas C é a presença de um centro de ligação de guani em sua subunidade a - nucleotídeos: GDP e GTP (Figuras 139, 145). Se o GTP estiver ligado à proteína C, isso corresponde ao seu estado ativado. Se o GDP estiver presente no centro de ligação de nucleotídeos, então esta forma corresponde ao estado inativo da proteína (Figura 79).
O acontecimento central na transmissão de um sinal do receptor, que é afectado pelo sinal primário, para a proteína G é que o receptor activado catalisa a troca de GDP ligado à proteína G por GTP presente no ambiente. Esta troca GDP/GTP na proteína G é acompanhada pela dissociação da molécula trimérica da proteína G em duas subunidades funcionais: a subunidade α contendo GTP e o complexo Py (Figuras 139, 145).
A seguir, uma dessas subunidades funcionais, que depende do tipo de sistema de sinalização, interage com a proteína efetora representada por uma enzima ou canal iônico. Como consequência, sua atividade catalítica ou condutância iônica muda de acordo, o que, por sua vez, leva a uma mudança na concentração citoplasmática do mensageiro secundário (ou íon) e, em última análise, inicia uma ou outra resposta celular.
As proteínas efetoras em sistemas de sinalização, como a proteína efetora da proteína GPCR-G, podem ser adenilato ciclase, que catalisa a síntese de AMPc a partir de ATP; fosfolipase C, que hidrolisa o fosfatidilinositol para formar DAT e 1P3; fosfodiesterase, que decompõe o cGMP em GMP; alguns tipos de canais de potássio e cálcio.
É importante que durante a transmissão do sinal na cascata do receptor da proteína G-proteína efetora, o sinal externo original possa ser amplificado (amplificado) muitas vezes. Isso ocorre porque uma molécula receptora, enquanto está no estado ativado (R*), consegue converter diversas moléculas de proteína G na forma ativada (G*).
Por exemplo, na cascata visual, a rodopsina^C^ecGMP-fosfodiesterase para cada molécula R* pode produzir várias centenas ou mesmo milhares de moléculas Gt*, o que significa que no primeiro estágio da cascata 7?*-»G* a o fator de amplificação do sinal externo é 10 2 -10 3 . Embora no próximo estágio da cascata (proteína efetora C*^) cada molécula G* interaja com apenas uma molécula da proteína efetora, o sinal aqui também é amplificado, pois para cada molécula de G* e, consequentemente, da proteína ativada proteína efetora, muitas aparecem (ou desaparecem) nas moléculas mensageiras secundárias do citoplasma. Assim, na cascata visual em seu segundo estágio, uma molécula de cGMP fosfodiesterase ativada é capaz de quebrar até 3.000 moléculas de cGMP por segundo, que serve como mensageiro secundário nas células fotorreceptoras.
O ganho total da cascata é igual ao produto dos ganhos em todos os estágios da cascata. O coeficiente de amplificação do sinal ao passar pela cascata pode atingir valores muito elevados: nas células visuais este é um valor da ordem de 10 5 -10 6 .
A cessação do estímulo externo é acompanhada pelo desligamento de todos os componentes do sistema de sinalização. Ao nível do receptor, isto é conseguido, em primeiro lugar, como resultado da dissociação do mensageiro primário do complexo com GPCR e, em segundo lugar, pela fosforilação dos receptores sob a acção de proteínas quinases especiais e subsequente ligação a um receptor modificado de um especial proteína (por exemplo, P-arrestina).
As proteínas G possuem atividade GTPase, ou seja, a capacidade de hidrolisar o GTP a elas associado em PIB, o que garante sua autoexclusão, ou seja, a transição G-GTP-e G-GDP. Como o estado de ativação da proteína efetora (on-off) depende diretamente do estado da proteína G, esta transição também significa o desligamento da proteína efetora e, conseqüentemente, a cessação da síntese (hidrólise) do mensageiro secundário ou o fechamento do canal iônico.
E, finalmente, para que a transição da célula ao seu estado original (antes do estímulo externo) seja concluída, mecanismos especiais restauram o nível inicial do mensageiro secundário ou cátion em seu citoplasma. Por exemplo, cAMP, cuja concentração citoplasmática aumenta durante a transmissão do sinal na cascata do receptor P-adrenérgico C 5 -proteína-adenilato ciclase, é então hidrolisado pela cAMP fosfodiesterase em AMP não cíclico (linear), que não possui as propriedades de um mensageiro secundário.
TRANSMISSÃO DE SINAL TRANSMEMBRANA. Uma propriedade importante das membranas é a capacidade de perceber e transmitir sinais do ambiente externo para a célula. O “reconhecimento” de moléculas sinalizadoras é realizado com a ajuda de proteínas receptoras incorporadas na membrana celular das células-alvo ou localizadas na célula.
Se o sinal for percebido pelos receptores de membrana, o esquema de transmissão de informações pode ser representado da seguinte forma:
interação do receptor com uma molécula sinalizadora (mensageiro primário);
ativação da enzima de membrana responsável pela formação do segundo mensageiro;
formação do mensageiro secundário cAMP, cGMP, IP3, DAT ou Ca 2+;
ativação por mediadores de proteínas específicas, principalmente proteínas quinases, que, por sua vez, por meio da fosforilação de enzimas, influenciam a atividade dos processos intracelulares.
Existem vários mecanismos de transferência transmembrana de informação: utilizando o sistema adenilato ciclase, sistema inositol fosfato, receptores catalíticos, receptores citoplasmáticos ou nucleares.
Organização estrutural e funcional das proteínas G
As proteínas G (proteínas de ligação ao GTP) são intermediários universais na transmissão de sinais dos receptores para as enzimas da membrana celular que catalisam a formação de segundos mensageiros do sinal hormonal. As proteínas G são oligômeros que consistem em subunidades α, β e γ.
Cada subunidade α na proteína G possui centros específicos:
vinculação de GTP ou PIB;
interações com o receptor;
ligação a subunidades βγ;
fosforilação pela proteína quinase C;
interação com a enzima adenilato ciclase ou fosfolipase C.
A estrutura das proteínas G carece de domínios α-helicoidais que abrangem a membrana. As proteínas G pertencem ao grupo das proteínas “ancoradas”.
Regulação da atividade da proteína G
Existe uma forma inativa da proteína G - o complexo αβγ-GDP e a forma ativada αβγ-GTP. A ativação da proteína G ocorre mediante interação com o complexo ativador-receptor; uma mudança na conformação da proteína G reduz a afinidade da subunidade α pela molécula GDP e aumenta-a pelo GTP. A substituição do GDP por GTP no sítio ativo da proteína G perturba a complementaridade entre as subunidades α-GTP e βγ. O receptor associado à molécula sinalizadora pode ativar um grande número de moléculas de proteína G, proporcionando assim a amplificação do sinal extracelular nesta fase.
A subunidade α da proteína G ativada (α-GTP) interage com uma proteína específica da membrana celular e modifica sua atividade. Tais proteínas podem ser as enzimas adenilato ciclase, fosfolipase C, cGMP fosfodiesterase, canais de Na+, canais de K+.
Arroz. 5-35. Ciclo de funcionamento da proteína G. Rs - receptor; G - hormônio; AC - adenilato ciclase.
O próximo estágio do ciclo de funcionamento da proteína G é a desfosforilação do GTP ligado à subunidade α, e a enzima que catalisa essa reação é a própria subunidade α.
A desfosforilação resulta na formação de um complexo α-GDP, que não é complementar a uma proteína de membrana específica (por exemplo, adenilato ciclase), mas tem alta afinidade por protômeros py. A proteína G retorna à sua forma inativa, αβγ-GDP. Com a subsequente ativação do receptor e substituição da molécula GDP por GTP, o ciclo se repete novamente. Assim, as subunidades α das proteínas G transportam um sinal estimulatório ou inibitório de um receptor que é ativado por um mensageiro primário (por exemplo, um hormônio) para uma enzima que catalisa a formação de um mensageiro secundário.
Algumas formas de proteínas quinases podem fosforilar as subunidades α das proteínas G. A subunidade α fosforilada não é complementar a uma proteína de membrana específica, como a adenilato ciclase ou a fosfolipase C e, portanto, não pode participar na transdução de sinal.
Adenilato-ciclase
A enzima adenilato ciclase, que catalisa a conversão de ATP em cAMP, é uma enzima chave no sistema de transdução de sinal da adenilato ciclase. A enzima pertence ao grupo das proteínas integrais da membrana celular e possui 12 domínios transmembrana. Fragmentos extracelulares da adenilato ciclase são glicosilados. Os domínios citoplasmáticos da adenilato ciclase possuem dois centros catalíticos responsáveis pela formação do AMPc, segundo mensageiro envolvido na regulação da atividade da enzima proteína quinase A.
A atividade da adenilato ciclase é influenciada por reguladores extracelulares e intracelulares. Os reguladores extracelulares (hormônios, eicosanóides, aminas biogênicas) realizam a regulação por meio de receptores específicos, que, por meio das subunidades α das proteínas G, transmitem sinais à adenilato ciclase. A subunidade α s (estimulante), ao interagir com a adenilato ciclase, ativa a enzima, a subunidade α (inibitória) inibe a enzima. Das 8 isoformas estudadas de adenilato ciclase, 4 são dependentes de Ca 2+ (ativadas por Ca 2+). A regulação da adenilato ciclase pelo cálcio intracelular permite que a célula integre as atividades dos dois principais segundos mensageiros cAMP e Ca2+.
Sistema adenilato ciclase
Com a participação do sistema adenilato ciclase, são realizados os efeitos de centenas de moléculas sinalizadoras de diversas naturezas - hormônios, neurotransmissores, eicosanóides.
O funcionamento do sistema de sinalização transmembrana é garantido por proteínas: receptor Rs da molécula sinalizadora, que ativa a adenilato ciclase, e receptor R i da molécula sinalizadora, que inibe a adenilato ciclase; proteínas estimuladoras de Gs e inibidoras de Gj-adenilato ciclase; enzimas adenilato ciclase (AC) e proteína quinase A (PKA).
A sequência de eventos que levam à ativação da adenilato ciclase:
a ligação de um ativador do sistema adenilato ciclase, por exemplo, o hormônio (G) ao receptor (R s), leva a uma mudança na conformação do receptor e a um aumento em sua afinidade pela proteína G s. Como resultado, forma-se o complexo [G][R][O-GDP];
a adição de [G][R] ao G-GDP reduz a afinidade da subunidade α da proteína G s pelo PIB e aumenta a afinidade pelo GTP. O PIB é substituído pelo GTP;
isso causa a dissociação do complexo. A subunidade α separada, associada a uma molécula de GTP, tem afinidade pela adenilato ciclase:
[G][R] → [G][R] + α-GTP + βγ;
a interação da subunidade α com a adenilato ciclase leva a uma mudança na conformação
enzima e sua ativação, a taxa de formação de AMPc a partir de ATP aumenta;
Mudanças conformacionais no complexo [α-GTP][AC] estimulam um aumento na atividade da GTP-fosfatase da subunidade α. Ocorre a reação de desfosforilação do GTP, e um dos produtos da reação, o fosfato inorgânico (Pi), é separado da subunidade α, e o complexo [α-GDP] é retido; a taxa de hidrólise determina o tempo de transmissão do sinal;
a formação de uma molécula GDP no centro ativo da subunidade α reduz sua afinidade pela adenilato ciclase, mas aumenta sua afinidade pelas subunidades βγ. A proteína G retorna à sua forma inativa;
se o receptor estiver associado a um ativador, por exemplo um hormônio, o ciclo de funcionamento da proteína G se repete.

Sistema adenilato ciclase
O componente intracelular mais importante das cascatas de sinalização são as proteínas G. Atualmente, são conhecidas cerca de 20 proteínas G diferentes. Por exemplo, Gs e Gi estimulam e inibem a adenilato ciclase, respectivamente; Gq ativa a fosfolipase C. Dentre as proteínas G das células sensoriais podemos destacar: fotorreceptor - Gt (transducina), olfativo - Golf e gustativo - Gg.
Em termos de estrutura, as proteínas G são heterotrímeros constituídos por três tipos de subunidades: a (alfa), b (beta) e g (gama), porém, em condições nativas, as subunidades beta e gama funcionam como um único complexo. Uma característica estrutural comum das proteínas G é a presença de sete hélices alfa transmembrana. A característica mais importante das proteínas G é a presença em sua subunidade α de um centro de ligação para nucleotídeos guanil: GDP (difosfato de guanisidina) e GTP (trifosfato de guanisidina). Se o GTP estiver ligado a uma proteína G, então isto corresponde ao seu estado ativado (G-GTP) ou, caso contrário, a proteína G está numa posição ativada. Se o GDP estiver presente no sítio de ligação do nucleotídeo, então esta forma (G-GDP) corresponde ao estado “desligado”. O ponto chave na transmissão do sinal do receptor (que foi afetado pelo sinal primário) para a proteína G é a catálise pelo receptor ativado da troca do GDP ligado à proteína G para o GTP presente no ambiente (GDP/GTP troca na proteína G).
Os receptores transmembrana fornecem as funções vitais básicas da célula: sinalização, transporte, proteção. O estudo do mecanismo de ação de diversos compostos biologicamente ativos, inclusive antivirais e antibacterianos, mostrou que os alvos mais específicos tanto para compostos medicinais quanto tóxicos (venenos) são os receptores celulares de humanos e microrganismos patogênicos. Uma parcela significativa dos receptores transmembrana é Receptores acoplados à proteína G (GPCRs), Cerca de metade de todos os medicamentos atualmente conhecidos atuam especificamente nos GPCRs. De todos os tipos de receptores de superfície celular, os GPCRs são os mais universais. Esses receptores ligam uma ampla gama de moléculas, desde pequenos neurotransmissores até grandes proteínas. Os GPCRs estão envolvidos em quase todos os processos vitais.
A diversidade de sinais transmitidos pelos GPCRs é garantida pelo acoplamento funcional de diferentes GPCRs entre si. Assim, é óbvio que o mecanismo mais universal de influência de compostos tóxicos e medicinais na célula se realiza através do efeito no aparelho receptor da célula, alterando sua conformação ou as principais características da conexão ligante-receptor, sua especificidade e reversibilidade.

Em 2018, o Jejum da Natividade começará em 28 de novembro. Durante este período, os crentes ortodoxos se preparam para celebrar o Natal...

Constituir família é o sonho da maioria das mulheres. Eles querem ter um marido amoroso e um monte de filhos. Mas nem sempre é um relacionamento...

Este artigo contém: a oração mais poderosa pelo divórcio - informações retiradas de todo o mundo, da rede eletrônica e...

Site de informações sobre ícones, orações, tradições ortodoxas. Oração pelos escândalos e brigas na família, com o marido, com os filhos...

O que seria do Ano Novo sem champanhe, tangerinas, Olivier, formol e o “arenque com casaco de pele” preferido de todos. Com o último...

Vamos preparar os ingredientes necessários para os biscoitos. A primeira coisa a fazer é colocar a água para ferver. Nós precisamos...

É possível cadastrar funcionário para o cargo de diretor financeiro - contador-chefe? O contador-chefe afirma...

O chefe de uma pequena empresa pode gerenciar facilmente o orçamento de forma independente. VERIFICADO! Se você conseguir...

A criação de novos projetos envolve uma justificação económica preliminar para a sua viabilidade, posterior...
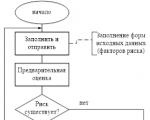
O reporte é gerado pelo RM, é acordado (aprovado) pelo Comité de Risco no âmbito do Conselho de Administração e transmitido a...

Na beira de um prado grande, muito grande, em uma longa folha de grama esmeralda vivia uma pequena joaninha. O pequeno de Deus...

Hoje em dia é bastante comum as pessoas se voltarem para as estrelas. Com a ajuda de um horóscopo uma pessoa pode descobrir...

Negócios ou amigável. Se você estava perseguindo um estranho, isso significa o seu nível de confiança no sexo feminino...

de acordo com o livro dos sonhos de Freud Se você sonhou que estava pescando, isso significa que na vida real dificilmente você consegue desligar...

Constituir família é o sonho da maioria das mulheres. Eles querem ter um marido amoroso e um monte de filhos. Mas nem sempre é...

Este artigo contém: a oração mais poderosa pelo divórcio - informações retiradas de todo o mundo, eletrônicas...