Azan e Iqamat (análise detalhada)
Ao ler o adhan, as mãos do muezzin devem tocar os lóbulos das orelhas e seu olhar deve estar direcionado para a Kaaba. Depois...
em biologia
"Química dos Hormônios"
Gonadotrofinas, FSH (hormônio folículo-estimulante, folitropina) e LH (hormônio luteinizante, lutropina) são glicoproteínas secretadas pelas células cianofílicas da adenohipófise (glândula pituitária anterior), sob a influência do fator de liberação hipotalâmico. Os órgãos-alvo são as gônadas.
O peso molecular do FSH é 33.000 daltons e do LH é 28.000 daltons. Ambos os hormônios têm uma estrutura dimérica, ou seja, consistem em uma cadeia alfa hormonalmente inespecífica (esta cadeia é intercambiável com FSH e LH) e uma cadeia beta que determina a especificidade hormonal. As cadeias são conectadas por ligações não covalentes. O alto teor de sacarídeos de até 16% também desempenha um papel importante na determinação da atividade biológica. Quando a casca sacarídica é removida da superfície do hormônio, este perde parcialmente sua atividade biológica, enquanto a atividade imunológica permanece inalterada (é possível a determinação por métodos imunológicos). A estrutura da cadeia alfa desses hormônios é muito semelhante à do hCG e do TSH. A cadeia alfa ou beta isolada praticamente não tem atividade biológica.
A estrutura molecular de ambos os hormônios é heterogênea, determinada pela idade e sexo e possivelmente induzida por esteróides sexuais. Ainda não foi estabelecido se esta heterogeneidade tem algum significado clínico.
Imagem 1. O mecanismo de ação dos proteohormônios (usando o exemplo do LH) na membrana plasmática da célula-alvo. P = receptor de membrana; B = proteína transmissora; A = adenilato ciclase; ATP = trifosfato de adenosina; AMP cíclico = 3,5-adenosina monofosfato cíclico
O FSH e o LH realizam sua atividade hormonal através de receptores na superfície da célula-alvo gonadal; nesse processo, o hormônio correspondente atua como mensageiro de primeira ordem. Então, no processo de uma cascata de reações sequenciais, o impulso hormonal se propaga na célula-alvo com a ajuda do AMP cíclico, um mensageiro de segunda ordem (ver Fig. 1). Em interação com o estradiol, o FSH induz seus receptores; Os receptores de LH também são induzidos pelo FSH.
Nas mulheres, o LH e o FSH têm um efeito sinérgico na biossíntese dos hormônios esteróides ovarianos. Os alvos do LH são as células da membrana ovariana e o corpo lúteo. Além disso, o LH provoca a ovulação e luteiniza as células da granulosa. O FSH controla o crescimento dos folículos até atingirem a maturidade e a prontidão para a ovulação e, a partir dos estágios iniciais da biossíntese de esteróides, a interação sinérgica de FSH e LH também estimula a biossíntese de estradiol pelas células da granulosa.
Nos homens, o LH estimula a biossíntese de testosterona nas células de Leydig dos testículos. O FSH controla o crescimento e a função dos túbulos seminíferos, especialmente a espermatogênese nas células de Sertoli.
Ao contrário das gonadotrofinas, a prolactina consiste em uma única cadeia peptídica composta por 198 resíduos de aminoácidos. Entre outras coisas, a estrutura espacial do hormônio é estabilizada por três pontes dissulfeto. A prolactina não contém resíduos sacarídeos, ou seja, não é uma glicoproteína. O peso molecular do hormônio é 22.000 dolton. Existem certas analogias estruturais com o hormônio do crescimento (somatotropina, hormônio do crescimento, hormônio do crescimento), bem como com o lactogênio placentário humano (HPL).
A prolactina que circula no sangue é caracterizada por polimorfismo molecular, ou seja, pode ser “pequeno”, “grande” e “muito grande”, e a imunogenicidade dessas formas é a mesma. Supõe-se que a prolactina “pequena” seja uma forma monomérica, e “grande” e “muito grande”, respectivamente, são tetraméricas. A prolactina “pequena” representa aproximadamente 80% da quantidade total do hormônio imunologicamente detectável no sangue, “grande” - 5-20% e “muito grande” - 0,5-5%. Além disso, o soro contém prolactina clivada, que é imunologicamente ativa e tem peso molecular de 8.000 a 16.000 daltons. Experimentos em animais mostraram um forte efeito mitogênico desta prolactina no tecido da glândula mamária.
Tal como as gonadotrofinas, a prolactina exerce os seus efeitos fisiológicos nas células-alvo através de receptores localizados na membrana. Juntamente com o estradiol, a prolactina nas mulheres afeta o crescimento e o funcionamento das glândulas mamárias e causa a lactação. Segundo alguns pesquisadores, a prolactina desempenha um papel na formação e função do corpo lúteo.
Nos homens, a função específica da prolactina não foi estabelecida.
A prolactina é sintetizada em células lactogênicas especializadas da glândula pituitária anterior; sua síntese e liberação estão sob a influência inibitória da limulação do hipotálamo. Além da glândula pituitária, a prolactina é produzida pela decídua (presença de prolactina no líquido amniótico) e pelo endométrio.
A luliberina (fator de liberação do hormônio luteinizante), também chamada de GnRH (fator de liberação de gonadotrofina) é um decapeptídeo de estrutura conhecida. A luliberina é sintetizada em células nervosas (neurônios) de certas áreas do hipotálamo ventral e mediobasal (Nucleus arcuatus, ventromedialis, periventricularis anterior, area preoptica suprachismatica), que são identificadas imuno-histoquimicamente. Através dos axônios das células nervosas, o hormônio é transportado para a eminência medial (Eminentia mediana), onde é liberado no sangue por um sistema portal especial que cobre o hipotálamo, o pedúnculo hipofisário e a adenohipófise. No lobo anterior da glândula pituitária, a luliberina estimula a síntese e liberação de LH e FSH através da ligação específica a receptores na membrana das células da adenohipófise. As variações no nível de gonadotrofinas nas mulheres, bem como as diferenças na proporção de FSH e LH dependendo da idade e das fases do ciclo menstrual, são provavelmente explicadas por mudanças no estado funcional das células gonadotrópicas da glândula pituitária anterior (variações no número de receptores de luliberina, que determina a sensibilidade dos gonadotrópicos a ela). A inibina também tem um efeito modelador neste mecanismo. A luliberina é catabolizada e inativada pelas endopeptidases da adenohipófise.
A inibina é um peptídeo com peso molecular de 23.000 daltons. Nas mulheres, o hormônio é encontrado no líquido folicular e nos homens é sintetizado nos túbulos seminíferos dos testículos. A inibina inibe seletivamente a liberação de FSH da glândula pituitária anterior.
Todos os hormônios esteróides são baseados na estrutura do ciclopentano-per-hidrofenantreno, também chamada de sistema de anel esterano, que consiste em quatro anéis fundidos saturados de hidrogênio, três dos quais têm seis membros e um tem cinco membros (ver Fig. 2). Ao substituir os átomos de hidrogênio por um grupo metil ou grupos oxo ou hidroxila contendo oxigênio, é alcançada uma variedade significativa de efeitos fisiológicos dos hormônios.
No corpo feminino, o local de síntese dos esteróides sexuais mais importantes (ou seja, estrogênios, gestágenos e andrógenos) são os ovários e o córtex adrenal, e durante a gravidez - a placenta. Os principais esteróides sexuais para o corpo masculino são os andrógenos, que são sintetizados nos testículos e, em pequenas quantidades, no córtex adrenal. Além do exposto, deve-se notar que o fornecedor do colesterol, cujos derivados são todos esteróides sexuais e hormônios adrenais, é o fígado.
Figura 2. Classificação dos hormônios esteróides por subclasses e seus representantes mais importantes
Os esteróides são lipofílicos, o que significa que têm baixa capacidade de dissolução em água. Portanto, no sangue, 95% dos hormônios esteróides estão ligados a proteínas de transporte específicas. O equilíbrio entre esteróides ligados e livres obedece à lei da ação das massas. Com a ajuda de proteínas de transporte, os hormônios são transportados para seus órgãos-alvo. Apenas esteróides livres e não ligados a proteínas são biologicamente ativos. A globulina de ligação a esteróides sexuais (SSBG) liga-se especificamente ao estradiol e aos andrógenos com baixa capacidade e alta afinidade, enquanto a globulina de ligação a corticosteróides (CSB) liga-se à progesterona e aos glicocorticóides. Além de sua função de transporte, proteínas séricas específicas de ligação a hormônios protegem os esteróides da inativação metabólica ao longo do caminho da glândula secretora até o órgão alvo. CCSG e CSG são glicoproteínas ácidas com peso molecular de 45.000 (CCSG) e 65.000 (CRSG). A síntese dos hormônios esteróides ocorre inteiramente no corpo humano (ver Fig. 3). A substância inicial, CoA, ativa o ácido acético, um composto de carbono a partir do qual o colesterol é formado como resultado de uma série de transformações, posterior condensação e ciclização. O colesterol é composto por 27 átomos de carbono e, entre outros componentes, é o principal componente da fase inicial da síntese dos hormônios esteróides.
Figura 3. Esquema de biossíntese de todas as subclasses de hormônios esteróides
Pelo encurtamento mediado enzimaticamente da cadeia lateral conectada à estrutura do esteróide na posição 17p em 6 átomos de carbono, a pregnenolona é formada. A pregnenolona é um esteróide C21, a partir do qual a progesterona é posteriormente formada pela oxidação do grupo hidroxila na posição 3 (3 no grupo oxo com o movimento simultâneo da ligação dupla de D 5 para D 4. A progesterona é o principal representante dos gestágenos. Ao mesmo tempo, a progesterona é o precursor de todos os glico e mineralcorticóides (ver Fig. 2), bem como dos andrógenos, que são derivados da progesterona como resultado da clivagem enzimática da cadeia lateral na posição 17 (3. Neste No processo, a 17-hidroxiprogesterona atua como um produto intermediário, o que significa que os andrógenos são esteróides C 19, cujo principal representante é a testosterona.
Ao final de uma cadeia sintética complexa, que consiste em diversas reações intermediárias, ocorre a síntese de estrogênios, cujo representante mais ativo é o estradiol. Os estrogênios são sintetizados a partir dos andrógenos removendo o grupo de canto metil C19 localizado entre os anéis A e B enquanto aromatiza ("fenolização") o anel A. O anel fenólico A é a característica mais característica de todos os estrogênios.
Todos os hormônios esteróides são destruídos no fígado por redução, adição de grupos hidroxila, seguida de conjugação com ácido glucurônico ou sulfatação. Como resultado, formam-se compostos solúveis em água que podem ser excretados na urina.
Ao contrário das gonadotrofinas, os hormônios esteróides exercem seu efeito biológico penetrando na célula ao longo de um gradiente de concentração e ligando-se a moléculas receptoras solúveis presentes no citosol, em vez de interagir com receptores de membrana (ver Fig. 4). Cada grupo de hormônios esteróides possui seus próprios receptores especiais. A conexão com o receptor correspondente é reversível e possui alta afinidade.
O complexo receptor-esteróide citoplasmático resultante é gradualmente ativado ou transformado e se move dentro do núcleo da célula - a localização do aparelho genético. Aqui, o complexo hormônio-receptor interage com um segmento molecular específico da cromatina, a chamada zona de interação nuclear, que corresponde a uma região molecular do complexo esteróide-receptor.
Como consequência da ligação do complexo receptor do hormônio esteroide ao aceitador nuclear, a dupla hélice do DNA é clivada, disponibilizando a informação genética.
Figura 4. O mecanismo de ação molecular dos hormônios esteróides na célula-alvo
Este segmento de DNA é transcrito em mRNA. Além disso, depois de separado, o mRNA entra no espaço citoplasmático. Na fase final, durante a interação do mRNA (como matriz) com o aparelho de síntese de proteínas ribossômicas, ocorre a neossíntese de proteínas.
O principal representante dos estrogênios é o estradiol (ver Fig. 5), que possui a maior atividade biológica. A estrona é formada a partir do estradiol por desidrogenação mediada enzimaticamente em C17 e não possui atividade biológica pronunciada (devido à sua baixa capacidade de ligação ao receptor e acúmulo insuficiente no núcleo da célula). Durante a gravidez, a estrona pode ser detectada no soro em concentrações crescentes. Nesse caso, o hormônio é sintetizado a partir do sulfato de desidroepiandrosterona (DHEA - S), formado no córtex adrenal fetal. Assim, a estrona é um dos indicadores que caracterizam o estado do feto.
Figura 5. Estrutura dos estrogênios biologicamente mais significativos
Outro grupo interessante de estrogênios são os estrogênios catecol, ou seja, esteróides, derivados do estradiol e da estrona e possuindo um grupo adicional na segunda posição do anel A. Isso os torna semelhantes às catecolaminas: adrenalina e norepinefrina. Os estrogênios catecol, entre outros locais, são sintetizados no hipotálamo, onde, segundo muitos pesquisadores, atuam como neurotransmissores, como as catecolaminas.
No corpo feminino, o estradiol é sintetizado nos ovários, na membrana e nas células da granulosa dos folículos. Durante a fase lútea do ciclo menstrual, o estradiol é sintetizado exclusivamente pelas células do revestimento folicular, enquanto as células da granulosa luteinizam e passam para a síntese de progesterona. Em caso de gravidez, a produção massiva de estrogênio é realizada pela placenta. Outros locais de síntese de estrogênio, principalmente estrona na pós-menopausa, incluem o córtex adrenal e o tecido adiposo periférico devido à sua capacidade de aromatizar andrógenos.
Não foram encontradas evidências clinicamente confiáveis da presença de secreção de estrogênio no corpo masculino.
Os órgãos-alvo do estrogênio são o útero, a vagina, a vulva, as trompas de falópio e as glândulas mamárias. Os hormônios desse grupo são responsáveis pelo desenvolvimento das características sexuais secundárias e determinam as características físicas e mentais características da mulher. Os estrogênios também causam o fechamento dos pontos de crescimento epifisário e, assim, participam da regulação do crescimento linear. Além disso, os estrogênios têm um efeito indutor sobre uma série de proteínas plasmáticas sintetizadas no fígado (por exemplo, SSSG, DSG, globulina ligadora de TSH-tiroxina, lipoproteínas, fatores de coagulação sanguínea). Nas células-alvo, o estradiol induz tanto seus receptores quanto os receptores de progesterona.
O principal representante deste grupo de hormônios é a progesterona (ver Fig. 2). Nas mulheres, a progesterona é secretada pelo corpo lúteo e, durante a gravidez, pela placenta.
Não há evidências clinicamente confiáveis da presença de síntese de progesterona em homens.
Para que a progesterona exiba seu efeito fisiológico no corpo feminino, é necessária uma exposição prévia ao estrogênio. Tal como acontece com o estradiol, a atividade da progesterona nas células-alvo é mediada por receptores específicos. A ativação desse grupo de receptores ocorre de forma semelhante ao mecanismo de ativação dos receptores de estrogênio.
O principal órgão-alvo da progesterona é o útero. O hormônio causa a transformação secretora do endométrio proliferativamente espessado, garantindo assim sua prontidão para a implantação de um óvulo fertilizado. Além disso, a progesterona tem uma importante função de controle no sistema gonadotrofina-esteróide gonadal e causa estimulação do centro térmico. Isto causa um aumento de 0,5 graus na temperatura corporal durante a fase lútea do ciclo menstrual após a ovulação.
A progesterona bloqueia a síntese de seus próprios receptores, assim como do bloqueador de estradiol. Na célula endometrial, a progesterona induz a 17 (5-hidroxiesteróide desidrogenase, que é uma enzima chave no metabolismo do estradiol e converte o estradiol em estrona praticamente inativa. Assim, através de seu mecanismo receptor, a progesterona evita a formação excessiva de estradiol endógeno na célula-alvo Este efeito da progesterona, em combinação com o seu efeito negativo nos receptores de estradiol, realizado através de um mecanismo receptor, pode ser definido como o efeito antiestrogênico da progesterona.
Os principais representantes dos andrógenos no corpo feminino são testosterona, androstenediol e sulfato de diidroepiandrosterona (DHEA-S) (ver Fig. 6).
Nos ovários, os andrógenos são secretados nas células da parte interna da casca do folículo, sua parede externa; androsterona e testosterona são sintetizadas a partir do colesterol sob a influência do LH.
Os andrógenos estimulam o crescimento dos pelos pubianos e axilares, aumentam a libido e afetam o tamanho do clitóris e dos grandes lábios. Os andrógenos modulam a produção de gonadotrofinas na glândula pituitária anterior. A hiperandrogenemia em mulheres leva à virilização e à fertilidade prejudicada. Isto torna importante a determinação de andrógenos no diagnóstico da infertilidade feminina.
Figura 6. Estrutura dos representantes biologicamente mais importantes dos andrógenos
Como todos os hormônios esteróides, a atividade dos andrógenos é mediada por receptores intracelulares. Porém, não é a testosterona que entra em contato com o receptor, mas a 5oc-diidrotestosterona (DHT), que é formada na célula-alvo pela redução enzimática da ligação dupla D 4 (devido à atividade da 5a-redutase).
No corpo masculino, os principais representantes dos andrógenos são a testosterona e a desidrotestosterona (DHT). Nos órgãos-alvo (próstata, vesículas seminais e pele), a testosterona atua como um pré-hormônio; isso significa que a testosterona, tendo atingido o órgão alvo, é convertida em desidrotestosterona pela 5a-redutase, e somente depois disso a desidrotestosterona exerce seu efeito biológico através do mecanismo receptor descrito acima. Em outros órgãos-alvo, como músculos e rins, o efeito dos andrógenos é mediado diretamente, ou seja, sem conversão enzimática. Atualmente, a comunidade científica considera a presença de um terceiro mecanismo de causa e efeito ao nível do hipotálamo e em outras áreas do cérebro sensíveis às influências endócrinas; a testosterona como tal não possui atividade hormonal própria, porém, após sofrer aromatização, é transformada em estradiol, neste caso adquirindo atividade biológica por meio da interação com receptores.
Em comparação com a testosterona, a atividade biológica de outros andrógenos, como androstenediona, diidroepiandrosterona, sulfato de desidroepiandrosterona, androsterona, epiandrosterona e etiocolanolona, é 5-20 vezes menor. A Tabela 2 mostra as concentrações normais dos andrógenos mais importantes no corpo masculino.
Concentrações normais dos andrógenos clinicamente mais significativos em homens.
Este andrógeno está presente em maior quantidade, mas assim como a testosterona, essencialmente não tem atividade androgênica. A fonte mais importante de testosterona são as células de Leydig dos testículos, que foram estabelecidas durante o exame de homens submetidos à castração. Apenas pequenas quantidades de testosterona são sintetizadas na periferia pela transformação de precursores. A testosterona apoia a espermatogênese, estimula o crescimento e o funcionamento das glândulas sexuais acessórias, bem como o desenvolvimento do pênis e do escroto. O hormônio tem efeito anabólico, principalmente nos ossos e músculos. Durante a puberdade, a presença de testosterona provoca o crescimento linear da laringe, o que leva ao aprofundamento da voz. Sob a influência da testosterona, forma-se um tipo de crescimento de cabelo masculino (um “triângulo” na parte superior do púbis, barba, cabelo no peito, queda de cabelo na testa e no topo da cabeça). Por seu efeito direto na medula óssea, além de ativar a síntese de eritropoietina nos rins, a testosterona estimula a eritropoiese. O hormônio também é necessário para manter a libido e a potência.
Com base em sua estrutura química, os hormônios, bem como outras substâncias biologicamente ativas de natureza regulatória (por exemplo, fatores de crescimento, interleucinas, interferons, quimiocinas, angiotensinas, Pg e vários outros) são divididos em peptídeos, esteróides, derivados de aminoácidos e ácido araquidônico.
Peptídeohormônios pertencem a substâncias polares que não podem penetrar diretamente nas membranas biológicas. Portanto, o mecanismo de exocitose é utilizado para sua secreção. Pela mesma razão, os receptores hormonais peptídicos são incorporados à membrana plasmática da célula-alvo, e a transmissão do sinal às estruturas intracelulares é realizada por segundos mensageiros.
TransmissãoARNm e a montagem da cadeia polipeptídica ocorre nas cisternas do retículo endoplasmático rugoso (ergastoplasma).
Pós-traduçãomodificação(por exemplo, glicosilação - ligação de moléculas de carboidratos à cadeia polipeptídica, resultando na formação de glicoproteínas) e a formação de vesículas secretoras ocorre no complexo de Golgi.
Exocitose: vesículas de membrana secretora contendo hormônios peptídicos (peptídeos, polipeptídeos, proteínas, glicoproteínas) fundem-se por dentro com o plasmalema da célula secretora; Como resultado, o conteúdo das vesículas aparece fora da célula. A exocitose estimula um aumento na concentração de Ca 2+ no citoplasma causado pela despolarização da membrana celular.
Esteroidehormônios: mineralocorticóides, glicocorticóides, andrógenos, estrogênios, progestágenos, calcitriol. Essas substâncias - derivados do colesterol - são apolares, por isso penetram livremente nas membranas biológicas. Por esse motivo, a secreção de hormônios esteróides ocorre sem a participação de vesículas secretoras. Pela mesma razão, os receptores para moléculas apolares estão localizados dentro da célula-alvo. Esses receptores são geralmente chamados de receptores nucleares.
O colesterol entra nas células vindo de fora.
A síntese de hormônios esteróides é um processo de múltiplas etapas que ocorre com a participação de dezenas de enzimas localizadas no retículo endoplasmático liso e nas mitocôndrias.
Derivadosaminoácidos- tirosina (hormônios da tireoide contendo iodo, norepinefrina, adrenalina e dopamina), histidina (histamina), triptofano (melatonina e serotonina)
Derivadostirosina
Moléculas apolares de tiroxina (T 4) e triiodotironina (T 3) são formadas por sua clivagem da tireoglobulina iodada (madura) nos fagolisossomos dos tireócitos e entram no sangue através do plasmalema da parte basal dos tireócitos. Seus receptores são nucleares.
As catecolaminas não penetram nas membranas; elas se acumulam nas vesículas secretoras e são secretadas pelas células por exocitose. Seus receptores estão embutidos na membrana plasmática das células-alvo.
Derivadohistidina- a histamina é um produto da descarboxilação da histidina. Os receptores de histamina estão incorporados na membrana plasmática das células-alvo.
Derivadostriptofano- melatonina ( N- acetil-5 ‑ metoxitriptamina) e serotonina (5 ‑ hidroxitriptamina). Seus receptores estão embutidos na membrana plasmática das células-alvo.
Derivadosaraquidônicoácidos(eicosanóides ou prostanóides). Eicosanóides (do grego. eikosi- vinte) consistem (como o ácido araquidônico) em 20 átomos de carbono. Estes incluem prostaglandinas (Pg), tromboxanos, prostaciclinas, leucotrienos, ácidos hidroxieicosotetraenóico (HETE, do inglês hidroxieicosatetraenóico) e epoxieicosotrienóico, bem como derivados desses ácidos. Todos os eicosanóides possuem atividade fisiológica alta e versátil, muitos deles funcionam apenas no interior da célula.
Araquidônicoácido- um ácido graxo mobilizado dos fosfolipídios da membrana celular pela fosfolipase A 2 . O ácido araquidônico também é formado durante a degradação do diacilglicerol. O ácido araquidônico é oxidado de três maneiras: com a ajuda de ciclooxigenases (formam-se a via da ciclooxigenase, PG, tromboxanos, prostaciclinas), lipoxigenase (via da lipoxigenase, formam-se leucotrienos, 5-HETE) e epoxigenase do ácido araquidônico (CYP2J2 da família do citocromo P450 , HETE e ácido epoxieicosotrienóico são formados).
Prostaglandinas. São conhecidos PgG 2 (precursor de PgH 2), PgH 2 (precursor de prostaciclinas e tromboxanos), PgI 2 (às vezes chamado de prostaciclina), PgD 2, PgE 2 e PgF 2 . Esses ácidos alifáticos endógenos biologicamente ativos regulam muitas funções: aumentam a permeabilidade da parede vascular, afetam a contratilidade das SMCs dos vasos sanguíneos e brônquios, alteram o limiar de sensibilidade à dor e estimulam a secreção de suco gástrico. Pg é usado para induzir o parto. O desenvolvimento de uma série de condições patológicas também está associado à ação da PGE (inflamação, asma brônquica, crescimento tumoral), e a PGE 2 é um poderoso pirogênio e modulador da metástase de células cancerígenas.
Prostaciclinas(incluindo PgI 2) são formados a partir de PgI 2, inibem a agregação plaquetária; causar relaxamento dos vasos sanguíneos SMC.
Tromboxanos- um grupo de compostos que afetam a agregação plaquetária também causa contração dos vasos sanguíneos do SMC.
Leucotrienos. Leucotrienos conhecidos A 4 (precursor de outros leucotrienos) B 4, C 4, D 4, E 4, F 4. Os leucotrienos afetam a atividade de muitos canais iônicos (diretamente ou por meio de proteínas quinases) e os processos de exocitose, causam contração das SMCs intestinais e dos vasos sanguíneos e são mediadores da inflamação.
HETE- os ácidos hidroxieicosotetraenóicos e seus derivados promovem a liberação de Ca 2+ dos estoques intracelulares de cálcio em monócitos e macrófagos, são fortes vasoconstritores e promovem a proliferação de diferentes tipos de células.
Epoxieicosotrienosácidos estimulam a liberação de Ca 2+ dos estoques intracelulares de cálcio, estimulam a troca transmembrana de Na + -H +, a proliferação celular e são vasodilatadores.
Também é formado a partir de fosfolipídios de membrana fatorativaçãoplaquetas(PAF), que é um dos espasmógenos mais poderosos.
Mecanismos ações hormônios sobre células–alvos
As interações intercelulares de informações implementadas no sistema endócrino fornecem a seguinte sequência de eventos:
hormônioreceptorCélulas alvo(segundointermediário)respondercélulas–alvos
Cada hormônio tem um efeito regulador na célula-alvo se, e somente se, se ligar como um ligante a uma proteína receptora específica dentro da célula-alvo.
Circulação V sangue. Os hormônios circulam no sangue livremente ou em combinação com proteínas que os ligam (T 4, T 3, hormônios esteróides, fatores de crescimento semelhantes à insulina, hormônio do crescimento). A ligação a essas proteínas aumenta significativamente a meia-vida dos hormônios. Assim, o T4 circula como parte do complexo por cerca de 1 semana, enquanto a meia-vida do T4 livre é de vários minutos.
Receptores e segundos mensageiros são discutidos no Capítulo 4, Interações célula-célula.
Hormônios(Grego Ορμόνη ) (grego) hormão- excitar, induzir) - produtos químicos sinalizadores biologicamente ativos secretados pelas glândulas endócrinas diretamente no corpo e que têm um efeito remoto, complexo e multifacetado no corpo como um todo ou em certos órgãos e tecidos alvo. Os hormônios servem como reguladores humorais (transmitidos pelo sangue) de certos processos em vários órgãos e sistemas.
Existem outras definições segundo as quais a interpretação do conceito de hormônio é mais ampla: “sinalizar substâncias químicas produzidas pelas células do corpo e que afetam as células de outras partes do corpo”. Esta definição parece preferível, uma vez que abrange muitas substâncias tradicionalmente classificadas como hormonas: hormonas animais que não possuem sistema circulatório (por exemplo, ecdisonas de lombrigas, etc.), hormonas de vertebrados que não são produzidas nas glândulas endócrinas (prostaglandinas, eritropoetina , etc.), bem como hormônios vegetais.
Inaugurado em 1902 por Starling e Bayliss.
Eles são usados no corpo para manter sua homeostase, bem como para regular muitas funções (crescimento, desenvolvimento, metabolismo, resposta a mudanças nas condições ambientais).
Todos os hormônios exercem seus efeitos no corpo ou em órgãos e sistemas individuais com a ajuda de receptores especiais para esses hormônios. Os receptores hormonais são divididos em 3 classes principais:
Todos os receptores são caracterizados pelo fenômeno de autorregulação da sensibilidade por meio de um mecanismo de feedback - quando o nível de um determinado hormônio está baixo, o número de receptores nos tecidos e sua sensibilidade a esse hormônio aumentam automaticamente de forma compensatória - processo denominado sensibilização (como bem como receptores de regulação positiva ou sensibilização (sensibilização). Por outro lado, quando o nível de um determinado hormônio está alto, ocorre uma diminuição compensatória automática no número de receptores nos tecidos e na sua sensibilidade a esse hormônio - um processo denominado dessensibilização (bem como regulação negativa ou dessensibilização) dos receptores.
Um aumento ou diminuição na produção de hormônios, bem como uma diminuição ou aumento na sensibilidade dos receptores hormonais e uma violação do transporte hormonal levam a doenças endócrinas.
Quando um hormônio no sangue atinge uma célula-alvo, ele interage com receptores específicos; os receptores “lêem a mensagem” do corpo e certas mudanças começam a ocorrer na célula. Cada hormônio específico corresponde exclusivamente a “seus próprios” receptores localizados em órgãos e tecidos específicos - somente quando o hormônio interage com eles é que se forma um complexo hormônio-receptor.
Os mecanismos de ação dos hormônios podem ser diferentes. Um dos grupos é composto por hormônios que se ligam a receptores localizados no interior das células – geralmente no citoplasma. Estes incluem hormônios com propriedades lipofílicas - por exemplo, hormônios esteróides (sexo, glicocorticóides e mineralocorticóides), bem como hormônios da tireoide. Sendo solúveis em gordura, esses hormônios penetram facilmente na membrana celular e começam a interagir com receptores no citoplasma ou núcleo. Eles são ligeiramente solúveis em água e, quando transportados pelo sangue, ligam-se a proteínas transportadoras.
Acredita-se que nesse grupo de hormônios o complexo hormônio-receptor atue como uma espécie de relé intracelular - formado na célula, passa a interagir com a cromatina, que está localizada no núcleo da célula e é composta por DNA e proteína, e assim acelera ou retarda o trabalho de certos genes. Ao influenciar seletivamente um gene específico, o hormônio altera a concentração do RNA e da proteína correspondentes e, ao mesmo tempo, corrige os processos metabólicos.
O resultado biológico de cada hormônio é muito específico. Embora os hormônios geralmente alterem menos de 1% das proteínas e do RNA na célula-alvo, isso é suficiente para obter o efeito fisiológico correspondente.
A maioria dos outros hormônios são caracterizados por três características:
O mecanismo de ação do complexo hormônio-receptor desses hormônios envolve necessariamente intermediários que induzem uma resposta celular. Os mais importantes desses mediadores são AMPc (adenosina monofosfato cíclico), trifosfato de inositol e íons cálcio.
Assim, em um ambiente desprovido de íons cálcio, ou em células com quantidade insuficiente deles, o efeito de muitos hormônios é enfraquecido; ao utilizar substâncias que aumentam a concentração intracelular de cálcio, ocorrem efeitos idênticos aos de alguns hormônios.
A participação dos íons cálcio como intermediário garante o efeito nas células de hormônios como a vasopressina e as catecolaminas.
No entanto, existem hormônios para os quais ainda não foi descoberto um mensageiro intracelular. Entre os mais famosos desses hormônios está a insulina, na qual foi proposto que o AMPc e o GMPc desempenhem o papel de mediador, assim como os íons de cálcio e até mesmo o peróxido de hidrogênio, mas ainda não há evidências convincentes a favor de qualquer substância. Muitos pesquisadores acreditam que, neste caso, os mediadores podem ser compostos químicos cuja estrutura é completamente diferente da estrutura dos mediadores já conhecidos pela ciência.
Tendo completado a sua tarefa, os hormônios são decompostos nas células-alvo ou no sangue, transportados para o fígado, onde são decompostos, ou finalmente eliminados do corpo principalmente através da urina (por exemplo, adrenalina).
Lista dos mais importantes:
| Estrutura | Nome | Redução | Lugar de síntese | Mecanismo de ação |
| triptamina | melatonina (N-acetil-5-metoxitriptamina) | Glândula pineal | ||
| triptamina | serotonina | 5-HT | células enterocromafins | |
| derivado de tirosina | tiroxina | T4 | tireoide | receptor nuclear |
| derivado de tirosina | triiodotironina | T3 | tireoide | receptor nuclear |
| adrenalina (epinefrina) | glândulas supra-renais | |||
| derivado de tirosina (catecolamina) | norepinefrina (norepinefrina) | glândulas supra-renais | ||
| derivado de tirosina (catecolamina) | dopamina | hipotálamo | ||
| peptídeo | hormônio anti-Mulleriano (substância inibitória Mulleriana) | AMG | Células de Sertoli | |
| peptídeo | adiponectina | tecido adiposo | ||
| peptídeo | hormônio adrenocorticotrófico (corticotropina) | ACTH | glândula pituitária anterior | acampamento |
| peptídeo | angiotensina, angiotensinogênio | fígado | IP3 | |
| peptídeo | hormônio antidiurético (vasopressina) | ADH | glândula pituitária posterior | |
| peptídeo | peptídeo natriurético atrial | ANF | coração | cGMP |
| peptídeo | polipeptídeo insulinotrópico dependente de glicose | GUI | Células K do duodeno e jejuno | |
| peptídeo | calcitonina | tireoide | acampamento | |
| peptídeo | hormônio liberador de corticotropina | AKGG | hipotálamo | acampamento |
| peptídeo | colecistocinina (pancreozimina) | CCK | I células do duodeno e jejuno | |
| peptídeo | eritropoetina | rins | ||
| peptídeo | hormônio folículo estimulante | FSH | glândula pituitária anterior | acampamento |
| peptídeo | gastrina | Células G gástricas | ||
| peptídeo | grelina (hormônio da fome) | Células épsilon das ilhotas pancreáticas, hipotálamo | ||
| peptídeo | glucagon (antagonista da insulina) | células alfa das ilhotas pancreáticas | acampamento | |
| peptídeo | hormônio liberador de gonadotrofina (luliberina) | GnRH | hipotálamo | IP3 |
| peptídeo | hormônio liberador de somatotropina (hormônio liberador de "hormônio do crescimento", somatocrinina) | GHRH | hipotálamo | IP3 |
| peptídeo | gonadotrofina coriónica humana | hCG, hCG | placenta | acampamento |
| peptídeo | lactogênio placentário | PL, HPL | placenta | |
| peptídeo | hormônio somatotrópico (hormônio do crescimento) | GH ou hGH | glândula pituitária anterior | |
| peptídeo | inibina | |||
| peptídeo | insulina | células beta das ilhotas pancreáticas | Tirosina quinase, IP 3 | |
| peptídeo | fator de crescimento semelhante à insulina (somatomedina) | IFR, IGF | Tirosina quinase | |
| peptídeo | leptina (hormônio da saciedade) | tecido adiposo | ||
| peptídeo | hormonio luteinizante | LH, LH | glândula pituitária anterior | acampamento |
| peptídeo | hormônio estimulador de melanócitos | MSG | glândula pituitária anterior | acampamento |
| peptídeo | neuropeptídeo Y | |||
| peptídeo | oxitocina | glândula pituitária posterior | IP3 | |
| peptídeo | polipeptídeo pancreático | PP | Células PP de ilhotas pancreáticas | |
| peptídeo | hormônio da paratireóide (hormônio da paratireóide) | PTH | corpo epitelial | acampamento |
| peptídeo | prolactina | glândula pituitária anterior | ||
| peptídeo | relaxina | |||
| peptídeo | secretina | SCT | intestino delgado superior | |
| peptídeo | somatostatina | SRIF | células delta das ilhotas pancreáticas, hipotálamo | |
| peptídeo | trombopoietina | fígado, rins | ||
| peptídeo | hormônio estimulador da tireoide | glândula pituitária anterior | acampamento | |
| peptídeo | hormônio liberador de tireotropina | TRH | hipotálamo | IP3 |
| glicocorticóide | cortisol | córtex adrenal | direto | |
| mineralocorticóide | aldosterona | córtex adrenal | direto | |
| esteróide sexual (andrógeno) | testosterona | testículos | receptor nuclear | |
| esteróide sexual (andrógeno) | desidroepiandrosterona | DHEA | córtex adrenal | receptor nuclear |
| esteróide sexual (andrógeno) | androstenediol | ovários, testículos | direto | |
| esteróide sexual (andrógeno) | diidrotestosterona | plural | direto | |
| esteróide sexual (estrogênio) | estradiol | aparelho folicular dos ovários, testículos | direto | |
| esteróide sexual (progesterona) | progesterona | corpo lúteo dos ovários | receptor nuclear | |
| esterol | calcitriol | rins | direto | |
| eicosanóide | prostaglandinas | fluido seminal | ||
| eicosanóide | leucotrienos | glóbulos brancos | ||
| eicosanóide | prostaciclina | endotélio | ||
| eicosanóide | tromboxano | plaquetas |
Os hormônios são produtos de secreção interna produzidos por glândulas especiais ou células individuais, liberados no sangue e distribuídos por todo o corpo, normalmente causando certo efeito biológico.
Os próprios hormônios não influenciam diretamente nenhuma reação celular. Somente ao entrar em contato com um determinado receptor, exclusivo dele, uma determinada reação é causada.
Os hormônios têm estruturas químicas diferentes. Isso faz com que eles tenham propriedades físicas diferentes. Os hormônios são divididos em solúveis em água e solúveis em gordura. Pertencer a uma dessas classes determina seu mecanismo de ação. Isso é explicado pelo fato de que os hormônios lipossolúveis podem penetrar facilmente na membrana celular, que consiste principalmente em uma bicamada lipídica, enquanto os hormônios solúveis em água não conseguem fazer isso. Nesse sentido, os receptores (R) para hormônios solúveis em água e gordura têm diferentes locais de localização (membrana e citoplasma). Ao entrar em contato com o receptor de membrana, o hormônio provoca uma cascata de reações na própria célula, mas não tem efeito sobre o material genético. O complexo do P citoplasmático e do hormônio pode atuar nos receptores nucleares e causar alterações no aparato genético, o que leva à síntese de novas proteínas. Vamos dar uma olhada nisso mais de perto.
O mecanismo de ação dos hormônios esteróides (solúveis em gordura)
I. Penetração do esteróide (C) na célula
II. Formação do complexo SR
Todos os hormônios esteróides P são proteínas globulares de aproximadamente o mesmo tamanho que se ligam aos hormônios com afinidade muito alta.
III. Transformação de CP em uma forma capaz de se ligar a aceitadores nucleares [CP]
Qualquer célula contém toda a informação genética. No entanto, com a especialização celular, a maior parte do DNA fica privada da capacidade de servir como modelo para a síntese de mRNA. Isto é conseguido dobrando-se em torno de proteínas histonas, levando à inibição da transcrição. A este respeito, o material genético de uma célula pode ser dividido em 3 tipos de DNA:
1. transcricionalmente inativo
2. constantemente expresso
3.induzido por hormônios ou outras moléculas sinalizadoras.
4. Ligação de [CP] ao aceitador de cromatina
Ressalta-se que esta etapa da ação C não foi totalmente estudada e apresenta uma série de questões controversas. Acredita-se que [CP] interage com regiões específicas do DNA de uma forma que permite que a RNA polimerase entre em contato com domínios específicos do DNA.
Uma experiência interessante mostrou que a meia-vida do mRNA aumenta quando estimulado por um hormônio. Isto leva a muitas contradições: não fica claro se um aumento na quantidade de mRNA indica que [CP] aumenta a taxa de transcrição ou aumenta a meia-vida do mRNA; ao mesmo tempo, o aumento da meia-vida do mRNA é explicado pela presença de um grande número de ribossomos em uma célula estimulada por hormônio, que estabilizam o mRNA ou outro efeito de [CP] desconhecido para nós no momento.
V. Iniciação seletiva de transcrição de mRNAs específicos; síntese coordenada de tRNA e rRNA
Pode-se presumir que o principal efeito da [CP] é afrouxar a cromatina condensada, o que leva à abertura do acesso às moléculas de RNA polimerase. Um aumento na quantidade de mRNA leva a um aumento na síntese de tRNA e rRNA.
VI. Processamento de RNAs primários
VII. Transporte de mRNA para o citoplasma
VIII. Síntese proteíca
IX. Modificação de proteína pós-tradução
No entanto, como mostram as pesquisas, este é o principal, mas não o único mecanismo possível de ação dos hormônios. Por exemplo, andrógenos e estrogênios causam um aumento no AMPc em algumas células, sugerindo que também existem receptores de membrana para hormônios esteróides. Isso mostra que os hormônios esteróides atuam em algumas células sensíveis, como os hormônios solúveis em água.
Intermediários secundários
Hormônios peptídicos, aminas e neurotransmissores, ao contrário dos esteróides, são compostos hidrofílicos e não são capazes de penetrar facilmente na membrana plasmática da célula. Portanto, eles interagem com receptores de membrana localizados na superfície celular. A interação hormônio-receptor inicia uma reação biológica altamente coordenada que pode envolver muitos componentes celulares, alguns dos quais estão localizados a uma distância considerável da membrana plasmática.
O cAMP é o primeiro composto que Sutherland, que o descobriu, chamou de “segundo mensageiro”, porque considerou o “primeiro mensageiro” o próprio hormônio, que causa a síntese intracelular do “segundo mensageiro”, que medeia o efeito biológico do primeiro.
Hoje, pelo menos 3 tipos de segundos mensageiros podem ser nomeados: 1) nucleotídeos cíclicos (cAMP e cGMP); 2) íons Ca e 3) metabólitos de fosfatidilinositol.
Com a ajuda de tais sistemas, um pequeno número de moléculas hormonais, ligando-se aos receptores, provoca a produção de um número muito maior de moléculas de segundos mensageiros, e estas, por sua vez, influenciam a atividade de um número ainda maior de moléculas proteicas. Assim, ocorre uma amplificação progressiva do sinal que ocorre inicialmente quando o hormônio se liga ao receptor.
De forma simplificada, a ação do hormônio através do AMPc pode ser representada da seguinte forma:
1. hormônio + receptor estereoespecífico
2. ativação da adenilato ciclase
3. Formação de AMPc
4. garantir reação coordenada por cAMP
Os cientistas modernos têm amplas oportunidades para estudar as mudanças no corpo humano que ocorrem ao experimentar certas emoções e sentimentos. Eles estudaram os níveis de hormônios e diversas substâncias que ocorrem quando uma pessoa se apaixona e descobriram que, independentemente de sexo, raça e idade, as mesmas transformações ocorrem em nível molecular. Como resultado, concluiu-se imediatamente que o amor é uma reação química.
Se isso é verdade ou não, se é possível fazer alguém se apaixonar por você, se é possível dar uma injeção ou tomar um comprimido para parar de sofrer por um amor não correspondido, nós vamos descobrir.
Os cientistas dizem que a Natureza colocou um significado racional por trás do sentimento brilhante que surge quando a luz de uma pessoa converge como uma cunha para outra. Isto é apenas uma poupança de energia para ambos os parceiros, especialmente para os homens. Os amantes, passando por certos estágios de sentimentos e relacionamentos, concentram todas as suas energias em uma pessoa com quem possam continuar sua linhagem familiar.
Acontece que para deixar descendentes não é preciso se espalhar entre muitas outras pessoas, ou seja, economiza-se energia. E o instinto de preservação da espécie é um estímulo “poderoso”, “escrito” nos nossos genes. É necessário para que a humanidade não se extinga.
A escolha de um objeto amoroso não depende da quantidade de hormônios. Fundamental é a experiência sócio-psicológica adquirida na primeira infância. Mas a força da excitação sexual, a reação aos estímulos sexuais, bem como as mudanças fisiológicas que ocorrem durante a excitação (fluxo sanguíneo para os órgãos genitais, secreção de uma secreção lubrificante neles e assim por diante) dependem do nível de hormônios.
Uma pessoa pode se apaixonar em menos de um segundo. Os cientistas descobriram que se um homem ou uma mulher vê um representante do sexo oposto que é extremamente atraente para eles, em uma fração de instante, 12 áreas diferentes de seu cérebro são excitadas ao mesmo tempo. Eles são responsáveis por liberar diversos “hormônios do amor”, garantindo respiração rápida, batimentos cardíacos, aumento da transpiração e outras reações.
Agora, em alguns países, estão testando a seleção de “verdadeiros entes queridos” usando análise molecular de urina. Aqui descobrimos as substâncias associadas à imunidade, que no homem e na mulher devem encaixar-se como a “chave de uma fechadura”. Os criadores prometem alta precisão do método, que atualmente é muito caro.
Os psicólogos distinguem entre os conceitos de “amor” e “paixão”. Apaixonar-se é um “estágio inicial” que pode não evoluir para amor. É caracterizada por uma cascata de reações químicas e mentais, como resultado das quais surgem “sintomas” especiais:
Ao realizar uma tomografia PET em uma pessoa apaixonada ao mesmo tempo em que lhe mostra uma fotografia do objeto da paixão, é possível notar um aumento de atividade nas mesmas áreas do cérebro que são ativadas ao consumir cocaína.
Houve outro estudo que também exigiu medir a atividade cerebral. Nesse caso, mostrar a fotografia de um ente querido enquanto infligia uma leve dor ao sujeito resultou em um efeito anestésico. Nas áreas do cérebro responsáveis pela percepção da estimulação dolorosa, o sinal era fraco. Quando lhes foi mostrada a foto de outra pessoa, não houve tal reação.
Quando uma pessoa está apaixonada, ela não vê as deficiências de sua amada, mas quando as vê e as aceita, já é uma transição para o próximo estágio - o amor. Entre eles geralmente existem vários outros estágios: quando cada um se irrita com as deficiências do outro, quando se reconciliam, e apenas o sétimo estágio é considerado sentimento real.
Assim, à questão de saber se o amor é um sentimento ou uma emoção, a resposta é que é um sentimento. Mas apaixonar-se pode ser chamado de conjunto (ou melhor, de fonte) de emoções.
Este período ocorre em 4 fases:
Vamos dar uma olhada mais de perto em quais hormônios formam o amor:

Este é um hormônio sexual masculino que também é produzido em pequenas quantidades nas mulheres. Suas principais funções são o desenvolvimento muscular, características de deposição de gordura subcutânea, bom funcionamento e formação dos órgãos genitais masculinos. Também afeta o interesse e a atração sexual de um homem por uma mulher e, se houver pouco disso, a partir da adolescência, esse homem não terá grande desejo de conhecer mulheres.
São hormônios femininos liberados na primeira fase do amor. São responsáveis pela formação do corpo de acordo com o tipo feminino, participam do ciclo menstrual, controlam o funcionamento do coração e a força dos ossos. Quando uma mulher vê um homem de quem gosta, seus níveis de estrogênio aumentam.
Estas são substâncias semelhantes a hormônios sintetizadas nas glândulas sudoríparas de uma pessoa de qualquer sexo. São eles que fazem você prestar atenção em um amante em potencial.
Quando uma pessoa que está em fase de busca por uma alma gêmea vê um “objeto” adequado, adrenalina e testosterona são liberadas em seu sangue. Sob a pele, a testosterona é convertida em androsterona, liberada pelo suor e absorvida pelas bactérias que vivem na pele. Cada pessoa possui um conjunto diferente de bactérias, então o cheiro do feromônio é diferente. Esse cheiro, embora não seja reconhecido conscientemente pela pessoa, é captado por um órgão especial localizado no nariz - o plexo vomeronasal. Analisa se o “código químico” do feromônio corresponde ao esperado e, em caso afirmativo, desencadeia a liberação de hormônios sexuais, dopamina e óxido nítrico. Se o “código” de um não corresponder ao do outro, não haverá atração apaixonada. Só pode haver relacionamentos de confiança, que com o tempo podem evoluir para amor.
Esta substância é produzida no cérebro e sua liberação no sangue provoca emoções positivas (por exemplo, uma sensação de satisfação durante o orgasmo). Se não for suficiente, a pessoa fica inquieta, ansiosa, fica deprimida e pode até desenvolver transtorno obsessivo-compulsivo. O excesso de dopamina inibe a produção de serotonina, por isso muitas pessoas ficam ansiosas logo no início de um relacionamento; Eles experimentam mudanças de humor, mas ao mesmo tempo tendem a pensar no objeto de sua paixão, alimentando-o assim.
À medida que a quantidade de serotonina aumenta, o interesse sexual diminui e a sensibilidade aos estímulos amorosos é perdida. Isto é típico da fase de separação.

Essas substâncias são produzidas nas glândulas supra-renais e possuem muitos receptores - em quase todos os órgãos internos. Eles são responsáveis por salvar vidas em situações estressantes, pois podem aumentar a frequência cardíaca, acelerar a quebra do glicogênio e das gorduras, das quais o corpo pode obter energia, e aumentar a pressão arterial. Além disso, são responsáveis pela excitação sexual e pelo orgasmo.
O principal hormônio responsável pelo amor é a dopamina. É produzido nas glândulas supra-renais e é um precursor de hormônios conhecidos como adrenalina e norepinefrina. Seu principal efeito é manter um nível suficiente de pressão arterial. Mas quando uma pessoa sente inconscientemente o “cheiro” de feromônios de uma pessoa do sexo oposto, a quantidade de dopamina aumenta acentuadamente. No amor não correspondido, a concentração desse hormônio passa por 2 fases. Na primeira fase, forma-se uma sensação de paixão; na segunda, diminuindo drasticamente, causa depressão grave.
A oxitocina como hormônio do amor surge já na fase dos relacionamentos de longo prazo, quando o primeiro amor já “acabou”. A oxitocina é responsável pela formação de confiança entre amantes; aumenta no sangue de mulheres e homens. Nos representantes do sexo forte, a oxitocina suprime o desejo de mudança e, quanto mais, mais o homem não gosta da atenção de outras mulheres. No entanto, ele mantém a capacidade de avaliar a atratividade feminina.
Nas mulheres, a oxitocina também é responsável pela sensação de orgasmo.
A rigor, não é um hormônio, mas sim um neurotransmissor que é liberado no sangue. Pela sua natureza química, provém da “família” das anfetaminas. É capaz de ativar a produção de adrenalina e vicia.
Quanto mais secreta a relação entre duas pessoas, mais ela se produz, mais “doce” é essa relação (você sente alegria, euforia, excitação). Vale ressaltar que existe muita dessa substância no chocolate amargo, mas quando consumido, é destruído tão rapidamente na boca, esôfago e estômago que a feniletilamina não chega ao intestino, de onde poderia entrar no sangue.
O fato de os hormônios serem responsáveis pelo estado de paixão é comprovado pelo fato de que no hipopituitarismo, doença em que a glândula pituitária não produz quantidade suficiente de hormônios de “comando”, inclusive hormônios sexuais, a pessoa não consegue se apaixonar. No entanto, seria muito errado considerar o amor apenas como reações químicas. Sim, surge sob a influência de hormônios, mas só se desenvolve sob a influência da personalidade de uma pessoa.
A principal tarefa desse hormônio é aumentar a pressão arterial, contraindo os vasos sanguíneos e reduzindo a quantidade de urina produzida. Mas também, trabalhando em conjunto com a oxitocina, promove a formação da unidade sexual e emocional entre os amantes.
Esses são os conhecidos hormônios da alegria. Eles são desenvolvidos já na fase de apego, causando sensação de felicidade nos relacionamentos de longo prazo, reduzindo as dores físicas e preservando a memória do tempo que passaram juntos. Atuam nos receptores opiáceos (os mesmos em que atua a morfina); são produzidos na glândula pituitária.
Você já sabe como são chamados os hormônios do amor, bem como seus efeitos. Isso significa que apresentá-los (ou adicioná-los) à comida ou à bebida pode despertar o amor em uma pessoa? Não, isso não significa isso.
Nos experimentos, foram administrados apenas testosterona, estrogênios e tirosina (aminoácido necessário para a síntese de adrenalina e norepinefrina), mas apenas com comprovada diminuição do nível desses hormônios. Ao mesmo tempo, as pessoas aumentaram o seu interesse geral pelo sexo oposto, mas não por qualquer pessoa específica.
Foi proposto o uso de feromônios como uma espécie de “poção do amor”. Eles são usados em perfumes e vendidos. Mas há armadilhas aqui:
Para atrair o amor de uma determinada pessoa, é melhor demonstrar-lhe a sua bondade (se for mulher) ou força moral e física (se for homem). Você pode persuadir essa pessoa a fazer algo em conjunto que seja próximo do extremo ou que exija experiências emocionais mútuas (por exemplo, ajudar em conjunto pessoas ou animais necessitados): em tais situações, o nível de adrenalina e norepinefrina, os hormônios do fase de atração, aumenta.
O professor de bioquímica Fisher, especialista que estudou a fundo quais substâncias são produzidas durante o amor e por quanto tempo existem no corpo, provou que o amor dura de 8 meses a 3 anos. É exatamente por quanto tempo existe a euforia e a vontade de estar sempre presente - para que o casal possa conceber e dar à luz um filho, e o pai possa estar por perto enquanto o bebê ainda está completamente indefeso e a mãe precisa de sua ajuda.
Há mais de 3 anos, existe um amor em que os amantes raramente se encontram ou são constantemente impedidos de se encontrarem ou de ficarem sozinhos. As relações harmoniosas também duram muito tempo (embora também vivam crises), onde já não é o amor que reina como um conjunto de reações químicas no cérebro, mas uma reação fixa que o casal trabalha e que se chama “verdadeira”. amor."
Embora, em geral, o amor seja causado por hormônios, esse sentimento não pode ser reduzido puramente à química. Os produtos químicos afetam a mente, que reage a cada produto químico de maneira diferente. A reação depende do estado do psiquismo, cuja natureza ainda é pouco conhecida.
No momento, está apenas comprovado que o psiquismo não depende totalmente do conteúdo inicial de diversas substâncias químicas no corpo (em geral) e no sistema nervoso (em particular). Os seguintes fatores influenciam como e por quanto tempo uma pessoa amará e se ela será propensa ao tipo de amor chamado “amor patológico” ou “amor viciante”:
São estes factores que têm a principal influência na natureza e duração do amor do seu escolhido (ou escolhido). A quantidade de hormônios, neste caso os hormônios sexuais masculinos (andrógenos), afeta apenas os homens. Então, se, devido à supressão dos testículos, glândulas supra-renais ou hipotálamo com glândula pituitária, um menino produziu poucos andrógenos, e isso não foi corrigido antes do final da puberdade, tal homem ficará mais “frio”, e será muito difícil prolongar seu amor. Em homens com quantidades normais ou ligeiramente elevadas de andrógenos, a principal coisa que influenciará o comportamento são as reações mentais.
Tudo o que pode ser usado para aumentar a duração do amor - mas não aquelas experiências e emoções vívidas que ocorrem apenas no seu primeiro estágio, mas um desejo sereno de estar mais frequentemente com o escolhido (escolhido) - pode ser feito usando o seguinte técnicas psicológicas. Eles influenciam apenas parcialmente os níveis de vários produtos químicos sintetizados no corpo:
Ao mesmo tempo, não se pode preencher totalmente o tempo livre do objeto da paixão: bastam 4 horas por dia, durante as quais vocês se comunicam e trabalham juntos.
Na medicina, existem vários anti-hormônios ou substâncias que bloqueiam os receptores dos hormônios liberados quando ocorre a paixão. Eles não se aplicam ao amor não correspondido.
Por exemplo, foram desenvolvidos bloqueadores dos receptores da vasopressina (Tolvaptan, Conivaptan e outros), mas são prescritos para o tratamento da insuficiência cardíaca e apresentam um efeito diurético em vez de um efeito “anti-amor”. Os bloqueadores adrenérgicos (Anaprilina, Metoprolol) tornam você insensível à adrenalina e à norepinefrina, mas só podem reduzir a pulsação ao olhar para o objeto da paixão, mas não conseguem se livrar dos sentimentos de amor.
É possível bloquear a dopamina (dopamina) - a chamada , mas os medicamentos que podem fazer isso causam efeitos colaterais desagradáveis. Eles são usados apenas para transtornos graves como transtornos depressivos, dissociais, depressivos de ansiedade e esquizofrenia. Mesmo que, como você pensa, a depressão se desenvolva num contexto de amor não correspondido, antes de adquirir antagonistas dos receptores de dopamina, o diagnóstico deve ser confirmado por um psiquiatra ou psicoterapeuta. Além disso, existem métodos mais suaves de tratamento de transtornos depressivos.
Nesta fase, tendo conhecimentos sobre o amor e a sua química, para acelerar o processo de “remoção da seta do Cupido”, os cientistas propõem recorrer ao tratamento com medicamentos serotoninérgicos, bem como antidepressivos envolvidos no transporte da serotonina. Isso se deve ao fato de que durante o amor a concentração de dopamina aumenta e, conseqüentemente, o nível de seu antagonista natural, que é a serotonina, diminui. O tratamento específico com serotonina sintética ainda está em fase de pesquisa. Enquanto os cientistas o injetaram em ratos de laboratório, após o que foi observado o seguinte resultado: o rato que recebeu serotonina rejeitou um parceiro permanente, mas começou a “buscar o amor” de todos os outros machos, indiscriminadamente.
Além disso, Mohammed Kabbaj e seus colegas estão atualmente desenvolvendo agentes que bloquearão simultaneamente os receptores de oxitocina e vasopressina. Os cientistas acreditam que isso ajudará a prevenir doenças e condições em pessoas associadas ao divórcio e ao amor infeliz.
Enquanto o novo medicamento não estiver pronto, se você não quiser ser tratado com antidepressivos, poderá usar os recursos químicos do seu próprio corpo para se livrar do amor infeliz:
Infelizmente, é impossível interromper a cascata de secreções de vários “hormônios do amor” para deixar de amar apenas por meios químicos. Você precisará parar de entrar em contato com o “objeto da paixão”, deletar seus contatos (se possível, dizer para ele não procurar reuniões ou ligar, pelo menos por um ano). Um bom método é descrever meticulosamente os traços negativos da pessoa que o rejeitou, focar neles e depois descrever todas as qualidades que você procura em seu amante.

Ao ler o adhan, as mãos do muezzin devem tocar os lóbulos das orelhas e seu olhar deve estar direcionado para a Kaaba. Depois...

Em 2018, o Jejum da Natividade começará em 28 de novembro. Durante este período, os crentes ortodoxos se preparam para celebrar o Natal...

Constituir família é o sonho da maioria das mulheres. Eles querem ter um marido amoroso e um monte de filhos. Mas nem sempre é um relacionamento...

Este artigo contém: a oração mais poderosa pelo divórcio - informações retiradas de todo o mundo, da rede eletrônica e...

Site de informações sobre ícones, orações, tradições ortodoxas. Oração pelos escândalos e brigas na família, com o marido, com os filhos...

O que seria do Ano Novo sem champanhe, tangerinas, Olivier, formol e o “arenque com casaco de pele” preferido de todos. Com o último...

Vamos preparar os ingredientes necessários para os biscoitos. A primeira coisa a fazer é colocar a água para ferver. Nós...

É possível cadastrar funcionário para o cargo de diretor financeiro - contador-chefe? O contador-chefe afirma...

O chefe de uma pequena empresa pode gerenciar facilmente o orçamento de forma independente. VERIFICADO! Se você conseguir...

A criação de novos projetos envolve uma justificação económica preliminar para a sua viabilidade, posterior...
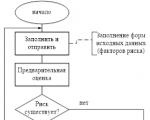
O reporte é gerado pelo RM, é acordado (aprovado) pelo Comité de Risco no âmbito do Conselho de Administração e transmitido a...

Na beira de um prado grande, muito grande, em uma longa folha de grama esmeralda vivia uma pequena joaninha. O pequeno de Deus...

Hoje em dia é bastante comum as pessoas se voltarem para as estrelas. Com a ajuda de um horóscopo uma pessoa pode descobrir...

Negócios ou amigável. Se você estava perseguindo um estranho, isso significa o seu nível de confiança no sexo feminino...

Em 2018, o Jejum da Natividade começará em 28 de novembro. Durante este período, os crentes ortodoxos se preparam para se encontrar...

Constituir família é o sonho da maioria das mulheres. Eles querem ter um marido amoroso e um monte de filhos. Mas nem sempre é...