Arenque sob um casaco de pele - uma receita clássica
O que seria do Ano Novo sem champanhe, tangerinas, Olivier, formol e o “arenque com casaco de pele” preferido de todos. Com o último...
1. Os procariontes não possuem membranas que limitam as organelas da célula bacteriana (núcleo, mitocôndrias, ribossomos) do citoplasma. Das membranas, existe apenas uma membrana citoplasmática.
2. O núcleo procariótico (nucleóide) possui estrutura fibrilar, o envelope nuclear está ausente.
3. Os procariontes não possuem mitocôndrias, cloroplastos e CG. EPS.
4. Os fragmentos redox estão localizados nos mesossomos (derivados da membrana citoplasmática)
5. Os procariontes não possuem mitose e se reproduzem por fissão binária.
6. Os procariontes têm um genoma haplóide.
7. Não há centro celular
8. Os movimentos intracelulares do citoplasma e os movimentos amebóides são atípicos para procariontes.
Características específicas do M/O
1. Tamanho pequeno, peso, volume e relativa simplicidade de estrutura.
2. Taxas de reprodução extremamente altas
3. Uma ampla variedade de maneiras de obter energia e através do metabolismo, ampla variedade produtos finais do metabolismo.
4. A capacidade de biodegradar quase todas as substâncias naturais e artificiais.
5. Extremamente alto grau adaptação como resultado de altas taxas de variabilidade.
6. População em massa e distribuição generalizada.
6. Estrutura e funções das formações superficiais de uma célula bacteriana. Cápsula. Métodos de detecção.
A célula bacteriana é cercada por uma membrana externa (Fig. 3.2), que consiste em uma cápsula, uma membrana semelhante a uma cápsula e uma parede celular. A capacidade da célula de perceber corantes de anilina (propriedades tintoriais) depende de sua composição. Dependendo da gravidade, as cápsulas são divididas em micro e macrocápsulas. Os primeiros são detectados apenas durante o exame microscópico eletrônico na forma de microfibrilas de mucopolissacarídeos, que estão intimamente adjacentes à parede celular. As macrocápsulas são uma camada mucosa pronunciada que cobre a parte externa da parede celular. Consiste em polissacarídeos e raramente polipeptídeos (por exemplo, na bactéria antraz). Via de regra, poucas espécies formam uma macrocápsula bactéria patogênica(toco de mococca, etc.) sob condições ambientais desfavoráveis, por exemplo no corpo de animais ou humanos. Porém, em algumas espécies (Klebsiella pneumoniae) a macrocápsula é constantemente encontrada.
O invólucro semelhante a uma cápsula é uma formação lipídica-polissacarídica que está relativamente fracamente associada à superfície celular, e como resultado, ao contrário da cápsula, pode ser liberada no meio ambiente.
A cápsula ou invólucro semelhante a cápsula pode ser revestido com exopolissacarídeos, que são formados a partir de carboidratos ambientais sob a ação de enzimas bacterianas. Ao mesmo tempo, glucanos e levanos garantem a adesão de bactérias a diferentes superfícies, muitas vezes lisas.
A cápsula carrega várias funções:
1. Protetor, protegendo a célula de condições adversas habitat,
2. adesivo, promovendo “aderência” à superfície (receptáculos) da célula hospedeira.
3. Frequentemente patogênico e propriedades antigênicas. Bactérias não patogênicas também podem formar uma macrocápsula, que aparentemente desempenha apenas uma função protetora.
7. Estrutura e funções da parede celular de bactérias gram-positivas e gram-negativas. Formas de bactérias com defeitos na parede celular.
Parede celular (CS) - um bioheteropolímero de composição química complexa que cobre toda a superfície de uma célula procariótica.
A base da parede celular é o peptidoglicano, que proporciona rigidez e elasticidade ao CS. A estrutura do peptidoglicano consiste em cadeias paralelas de polissacarídeos (glicanos) que consistem em unidades alternadas [\"-acetil1 lnjosamina E Ácido N-acetilmurâmico Um tripéptido está ligado covalentemente a cada resíduo de ácido N-acetidmurâmico
Diferenças entre bactérias Gram+ e Gram-.
| Grupo | 1" aríete -*-_^____________________ | grama - |
| Coloração de Gram | roxo | rosa |
| Espessura do policial | 20-60nm | 10-20nm |
| % conteúdo lipídico | 1,6% | 22,6% |
| Estrutura do peptidoglicano | Os peptídeos peptidoglicanos estão ligados através de uma ponte peptidil de 5 resíduos de glicina. | Os ácidos acetilmurâmicos de cada cadeia de glicano são conectados através de dois tetrapeptídeos do mesmo tipo |
| % de conteúdo de peptidoglicano | 40-90% Multicamadas | 5-10% Camada única |
| Presença de ácidos teicóicos | Disponível | Nenhum |
| Características do isolamento enzimático | As enzimas são liberadas de forma inconsistente no meio ambiente | As enzimas são secretadas no espaço periplasmático localizado entre o CS e o CM |
| Representantes | Todos os cocos patogênicos, exceto gonococos e meningococos, bacilos, iclostrdia | Enterobacteriaceae, Vibrio, Treponema |
Funções do KS:
1. Dá à célula uma certa forma.
2. Protege das influências ambientais
3. Carrega uma variedade de receptores na superfície aos quais se fixam alguns fagos, colishins e compostos químicos.
4. Através do CS eles entram na célula nutrientes e produtos metabólicos são liberados
5. Restringe a alta pressão osmótica intracelular.
CS Gram-bact. É representado por uma membrana externa de três camadas (peptidoglicano + lipopolissacarídeo + lipoproteínas). Algumas proteínas (norinas), penetrando na membrana externa, formam poros por onde passam substâncias hidrofílicas com baixo peso molecular.
O peptidoglicano é alvo da ação de alguns antibióticos (penicilina) e enzimas (lisozima). A penicilina interrompe a formação de ligações tetrapeptídicas, a lisozima destrói as ligações glicosídicas entre o ácido murâmico e a acetilglucosamina.
Quando a penicilina atua em um tanque em crescimento. cultura é formada formas sem casca bactérias:
1 Os protoplastos são completamente desprovidos de CS.
2. Esferoplastos - parcialmente desprovidos de CS
Tanto os protoplastos quanto os esferoplastos sofrem plasmólise em um ambiente isotônico, mas em um ambiente de trigo exibem fraca atividade metabólica! perder a capacidade de reprodução.
3.Formas L - total ou parcialmente desprovidas de CS, retêm a capacidade de reprodução.
a) estável - capaz de reverter à sua forma original.
b) instável - incapaz de reversão
8. Estruturas citopasmáticas de bactérias, funções, métodos de detecção. Micróbios ácido-resistentes. Método de coloração.
Flagelos. Os flagelos estão localizados na superfície de várias células bacterianas (Fig. 3.5). Contêm a proteína flagelina, que em sua estrutura pertence às proteínas contráteis do tipo miosina. Os flagelos estão ligados corpo basal, consistindo em um sistema de vários discos embutidos na membrana citoplasmática e no CS. O número e a localização dos flagelos variam entre as diferentes bactérias.
Monotrichs possuem apenas um flagelo em um dos pólos celulares,
lofotriquia - um feixe de flagelos,
anfitríquio - os flagelos estão localizados em ambos os pólos da célula,
peritríquio - Em toda a sua superfície.
Os flagelos têm propriedades antigênicas.
Bebido- finos filamentos ocos de natureza proteica, com 0,3-10 mícrons de comprimento e 10 nm de espessura, cobrindo a superfície das células bacterianas. Ao contrário dos flagelos, eles não desempenham função locomotora. De acordo com a sua finalidade funcional são divididos em vários tipos.
Bebi 1 tipo geral causar a ligação ou adesão de bactérias a certas células do corpo hospedeiro. Seu número é grande - de várias centenas a vários milhares por célula bacteriana. A adesão é o estágio inicial de qualquer processo infeccioso.
Bebi 2 tipos(sinônimo: conjugativo, ou sexual, bebeu - pili sexual) participam da conjugação de bactérias, o que garante a transferência de parte do material genético da célula doadora para a célula receptora. Eles estão disponíveis apenas em bactérias doadoras em quantidades limitadas (1-4 por célula).
Membrana citoplasmática (CM)é um componente estrutural vital da célula bacteriana. Limita o protoplasto, localizado diretamente abaixo da parede celular. Quimicamente, CM é uma lipoproteína composta por 15-30% de lipídios e 50-70% de proteínas. Além disso, contém cerca de 2 a 5% de carboidratos e uma pequena quantidade de RNA. Os lipídios da membrana consistem principalmente em lipídios neutros e fosfolipídios. Algumas bactérias contêm glicolipídios e os micoplasmas contêm esteróis.
A composição lipídica das membranas é qualitativa e quantitativamente variável. Para o mesmo tipo de bactéria, varia dependendo das condições de seu cultivo em meio nutriente e da idade da cultura. Tipos diferentes as bactérias diferem umas das outras na composição lipídica de suas membranas.
As proteínas da membrana são divididas em estruturais e funcionais. Estes últimos incluem enzimas envolvidas na biossíntese de vários componentes do CM, que ocorre na superfície do CM, bem como enzimas redox, permeases, etc.
O CM é uma estrutura complexamente organizada que consiste em três camadas, que são reveladas por microscopia eletrônica. A dupla camada fosfolipídica é permeada por globulinas, que garantem o transporte de substâncias para o interior da célula bacteriana.
CMs desempenham funções vitais, cuja violação leva à morte da célula bacteriana. Estes incluem, em primeiro lugar, a regulação da entrada de metabólitos e íons na célula, a participação
no metabolismo, na replicação do DNA e em várias bactérias na esporulação, etc.
Mesossomos são derivados de CM. Eles têm estruturas diferentes bactérias diferentes, localizado em partes diferentes células na forma de membranas concêntricas, ou vesículas, tubos, ou na forma de uma alça, característica principalmente de bactérias gram-negativas. Os mesossomos estão associados ao nucleóide. Eles estão envolvidos na divisão celular e na esporulação.
O citoplasma em procariontes, assim como em eucariotos, é um sistema coloidal complexo que consiste em água (cerca de 75%), compostos minerais, proteínas, RNA e DNA, que fazem parte de organelas nucleóides, ribossomos, mesossomos e inclusões.
Nucleóideé equivalente ao núcleo dos eucariotos, embora difira dele em sua estrutura e composição química. Não possui membrana nuclear, não contém cromossomos e não se divide por mitose. O nucleóide não contém as principais proteínas - histonas. As únicas exceções são algumas bactérias. Ele contém uma molécula de DNA de fita dupla e não é um grande número de RNA e proteínas. Uma molécula de DNA com peso molecular de (2-3) x 10 9 é uma estrutura de anel fechado na qual todas as informações hereditárias da célula são codificadas, ou seja, genoma celular. Por analogia com os cromossomos eucarióticos, o DNA bacteriano é frequentemente chamado de cromossomo. Deve-se lembrar que é apresentado em uma célula em singular, uma vez que as bactérias são haplóides. No entanto, antes da divisão celular o número de nucleóides duplica e durante a divisão aumenta para 4 ou mais.
Unidade da estrutura celular.
O conteúdo de qualquer célula é separado de ambiente externo estrutura especial - membrana plasmática (plasmalema). Este isolamento permite criar um ambiente muito especial dentro da célula, diferente do que a rodeia. Portanto, processos que não ocorrem em nenhum outro lugar podem ocorrer na célula; eles são chamados Processos da vida.
O ambiente interno de uma célula viva, delimitado pela membrana plasmática, é denominado citoplasma. Inclui hialoplasma(substância básica transparente) e organelas celulares, bem como várias estruturas não permanentes - inclusões. Organelas que estão presentes em qualquer célula também incluem ribossomos, onde isso acontece síntese proteíca.
A estrutura das células eucarióticas.
Eucariotos- São organismos cujas células possuem núcleo. Essencial- esta é a própria organela da célula eucariótica na qual é armazenada a informação hereditária registrada nos cromossomos e a partir da qual a informação hereditária é transcrita. Cromossomaé uma molécula de DNA integrada a proteínas. O núcleo contém nucléolo- o local onde se formam outras organelas importantes envolvidas na síntese de proteínas - ribossomos. Mas os ribossomos são formados apenas no núcleo e funcionam (ou seja, sintetizam proteínas) no citoplasma. Alguns deles estão livres no citoplasma e alguns estão ligados às membranas, formando uma rede, que é chamada endoplasmático.
Ribossomos- organelas não membranares.
Retículo endoplasmáticoé uma rede de túbulos delimitados por membrana. Existem dois tipos: liso e granular. Os ribossomos estão localizados nas membranas do retículo endoplasmático granular, de modo que as proteínas são sintetizadas e transportadas para lá. E o retículo endoplasmático liso é o local de síntese e transporte de carboidratos e lipídios. Não há ribossomos nele.
A síntese de proteínas, carboidratos e gorduras requer energia, que é produzida na célula eucariótica pelas “estações de energia” da célula - mitocôndria.
Mitocôndria- organelas de membrana dupla nas quais ocorre o processo de respiração celular. Os compostos orgânicos são oxidados nas membranas mitocondriais e a energia química é acumulada na forma de moléculas de energia especiais. (ATP).
Há também um local na célula onde os compostos orgânicos podem se acumular e de onde podem ser transportados - este é Aparelho de Golgi, sistema de sacos de membrana plana. Está envolvido no transporte de proteínas, lipídios e carboidratos. O aparelho de Golgi também produz organelas para digestão intracelular - lisossomos.
Lisossomos- organelas de membrana única, características das células animais, contêm enzimas que podem quebrar proteínas, carboidratos, ácidos nucleicos, lipídios.
Uma célula pode conter organelas que não possuem estrutura de membrana, como ribossomos e um citoesqueleto.
Citoesqueleto- este é um músculo-esquelético sistema celular, inclui microfilamentos, cílios, flagelos, um centro celular que produz microtúbulos e centríolos.
Existem organelas características apenas das células vegetais - plastídios. Existem: cloroplastos, cromoplastos e leucoplastos. O processo de fotossíntese ocorre nos cloroplastos.
Nas células vegetais também vacúolos- resíduos da célula, que são reservatórios de água e compostos nela dissolvidos. EM organismos eucarióticos incluem plantas, animais e fungos.
A estrutura das células procarióticas.
Procariontes- organismos unicelulares cujas células não possuem núcleo.
As células procarióticas são pequenas e armazenam material genético na forma de uma molécula circular de DNA (nucleóide). Os organismos procarióticos incluem bactérias e cianobactérias, anteriormente chamadas de algas verde-azuladas.
Se o processo de respiração aeróbica ocorre em procariontes, então saliências especiais da membrana plasmática são usadas para isso - mesossomos. Se as bactérias são fotossintéticas, então o processo de fotossíntese ocorre nas membranas fotossintéticas - tilacóides.
A síntese de proteínas em procariontes ocorre em ribossomos. As células procarióticas possuem poucas organelas.
Hipóteses da origem das organelas das células eucarióticas.
As células procarióticas apareceram na Terra antes das células eucarióticas.
1) hipótese simbiótica explica o mecanismo de surgimento de algumas organelas da célula eucariótica - mitocôndrias e plastídios fotossintéticos.
2) Hipótese de intussuscepção- afirma que a origem da célula eucariótica vem do fato de a forma ancestral ser um procarioto aeróbico. As organelas nele contidas surgiram como resultado da invaginação e descolamento de partes da casca, seguida de especialização funcional no núcleo, mitocôndrias, cloroplastos e outras organelas.
A característica mais importante e fundamental das células eucarióticas está associada à localização do aparelho genético na célula. O aparato genético de todos os eucariotos está localizado no núcleo e é protegido pelo envelope nuclear (em grego, “eucarioto” significa ter núcleo). O DNA dos eucariotos é linear (nos procariontes, o DNA é circular e está localizado em uma região especial da célula - o nucleóide, que não é separado por uma membrana do resto do citoplasma). Está associado a proteínas histonas e outras proteínas cromossômicas que as bactérias não possuem.
No ciclo de vida dos eucariotos, geralmente existem duas fases nucleares (haplofase e diplofase). A primeira fase é caracterizada por um conjunto haplóide (único) de cromossomos, então, fundindo-se, duas células haplóides (ou dois núcleos) formam uma célula diplóide (núcleo) contendo um conjunto duplo (diplóide) de cromossomos. Às vezes, durante a próxima divisão, e mais frequentemente após várias divisões, a célula torna-se novamente haplóide. Tal ciclo de vida e, em geral, a diploide não são típicos dos procariontes.
A terceira, talvez a mais interessante diferença, é a presença nas células eucarióticas de organelas especiais que possuem seu próprio aparato genético, se reproduzem por divisão e são circundadas por uma membrana. Essas organelas são mitocôndrias e plastídios. Em sua estrutura e atividade vital, eles são surpreendentemente semelhantes às bactérias. Esta circunstância levou os cientistas modernos a acreditar que tais organismos são descendentes de bactérias que mantiveram uma relação simbiótica com os eucariotos. Os procariontes são caracterizados por um pequeno número de organelas e nenhuma delas é cercada por uma membrana dupla. As células procarióticas não possuem retículo endoplasmático, aparelho de Golgi ou lisossomos.
Outra diferença importante entre procariontes e eucariotos é a presença de endocitose em eucariotos, incluindo fagocitose em muitos grupos. A fagocitose (literalmente “comer por uma célula”) é a capacidade das células eucarióticas de capturar, encerrar em uma vesícula de membrana e digerir uma ampla variedade de partículas sólidas. Este processo fornece uma importante função protetora no corpo. Foi descoberto pela primeira vez por II Mechnikov em estrelas do mar. O aparecimento de fagocitose em eucariotos está provavelmente associado ao tamanho médio (mais sobre diferenças de tamanho está escrito abaixo). Os tamanhos das células procarióticas são desproporcionalmente menores e, portanto, no processo desenvolvimento evolutivo eucariontes, eles tinham o problema de fornecer ao corpo uma grande quantidade de alimentos. Como resultado, os primeiros predadores reais e móveis aparecem entre os eucariotos.
A maioria das bactérias possui uma parede celular diferente da eucariótica (nem todos os eucariotos a possuem). Nos procariontes, é uma estrutura durável que consiste principalmente em mureína (em archaea, pseudomureína). A estrutura da mureína é tal que cada célula é cercada por um saco de malha especial, que é uma molécula enorme. Entre os eucariotos, muitos protistas, fungos e plantas possuem parede celular. Nos fungos consiste em quitina e glucanos, nas plantas inferiores consiste em celulose e glicoproteínas, nas diatomáceas sintetizam uma parede celular a partir de ácidos silícicos, nas plantas superiores consiste em celulose, hemicelulose e pectina. Aparentemente, para células eucarióticas maiores tornou-se impossível criar uma parede celular de alta resistência a partir de uma única molécula. Esta circunstância poderia forçar os eucariontes a usar materiais diferentes para a parede celular. Outra explicação é que o ancestral comum dos eucariotos perdeu sua parede celular devido à transição para a predação, e então os genes responsáveis pela síntese da mureína também foram perdidos. Quando alguns eucariontes retornaram à nutrição osmotrófica, a parede celular apareceu novamente, mas em uma base bioquímica diferente.
O metabolismo das bactérias também é diverso. Em geral, existem quatro tipos de nutrição e todos são encontrados entre as bactérias. Estes são fotoautotróficos, fotoheterotróficos, quimioautotróficos, quimioheterotróficos (fototróficos usam energia luz solar, quimiotrófico usa energia química). Os eucariotos sintetizam eles próprios a energia da luz solar ou usam energia pronta dessa origem. Isto pode ser devido ao surgimento de predadores entre os eucariotos, para os quais a necessidade de sintetizar energia desapareceu.
Outra diferença é a estrutura dos flagelos. Nas bactérias, eles são finos - apenas 15-20 nm de diâmetro. Estes são filamentos ocos feitos da proteína flagelina. A estrutura dos flagelos eucarióticos é muito mais complexa. Eles são um crescimento celular cercado por uma membrana e contêm um citoesqueleto (axonema) de nove pares de microtúbulos periféricos e dois microtúbulos no centro. Ao contrário dos flagelos procarióticos giratórios, os flagelos eucarióticos dobram-se ou contorcem-se. Os dois grupos de organismos que estamos considerando, como já mencionado, são muito diferentes nos seus tamanhos médios. O diâmetro de uma célula procariótica é geralmente de 0,5 a 10 mícrons, enquanto o mesmo valor para eucariotos é de 10 a 100 mícrons. O volume dessa célula é 1.000-10.000 vezes maior que o de uma célula procariótica. Os procariontes possuem ribossomos pequenos (tipo 70S). Os eucariotos possuem ribossomos maiores (tipo 80S).
Aparentemente, a época de surgimento desses grupos também difere. Os primeiros procariontes surgiram no processo de evolução há cerca de 3,5 bilhões de anos, a partir deles evoluíram organismos eucarióticos há cerca de 1,2 bilhão de anos.
Com base na sua estrutura celular, os organismos vivos são divididos em procarioto E eucariontes. As células de ambos estão cercadas membrana de plasma, fora dos quais, em muitos casos, há parede celular. Dentro da célula há um semilíquido citoplasma. No entanto, as células procarióticas são muito mais simples que as células eucarióticas.
Material genético básico procarioto (do grego sobre– antes e caryon– núcleo) está localizado no citoplasma na forma de uma molécula circular de DNA. Esta molécula ( nucleóide) não é circundado pelo envelope nuclear característico dos eucariotos e está ligado à membrana plasmática (Fig. 1). Assim, os procariontes não possuem núcleo formado. Além do nucleóide, uma pequena molécula circular de DNA chamada plasmídeo. Os plasmídeos podem mover-se de uma célula para outra e integrar-se na molécula principal de DNA.
Alguns procariontes possuem extensões de membrana plasmática: mesossomos, tilacóides lamelares, cromatóforos. Eles contêm enzimas envolvidas nos processos de fotossíntese e respiração. Além disso, os mesossomos estão associados à síntese de DNA e à secreção de proteínas.
As células procarióticas são pequenas em tamanho e seu diâmetro é de 0,3–5 μm. Na parte externa da membrana plasmática de todos os procariontes (com exceção dos micoplasmas) está parede celular. É composto por complexos de proteínas e oligossacarídeos, dispostos em camadas, protege a célula e mantém sua forma. É separado da membrana plasmática por um pequeno espaço intermembranar.
Apenas organelas não membranares são encontradas no citoplasma dos procariontes ribossomos. A estrutura dos ribossomos de procariontes e eucariotos é semelhante, porém, os ribossomos dos procariontes são menores e não estão ligados à membrana, mas estão localizados diretamente no citoplasma.

Muitos procariontes são móveis e podem nadar ou planar usando flagelos.
Os procariontes geralmente se reproduzem dividindo-se em dois ( binário). A divisão é precedida por um estágio muito curto de duplicação ou replicação cromossômica. Portanto, os procariontes são organismos haplóides.
Os procariontes incluem bactérias e algas verde-azuladas, ou cianobactérias. Os procariontes surgiram na Terra há cerca de 3,5 bilhões de anos e foram provavelmente as primeiras formas de vida celular, dando origem aos procariontes e eucariontes modernos.
Eucariotos (do grego ai credo- verdadeiro, caryon- núcleo) ao contrário dos procariontes, têm um núcleo formado rodeado envelope nuclear– membrana de dupla camada. As moléculas de DNA encontradas no núcleo são abertas (moléculas lineares). Além do núcleo, parte da informação genética está contida no DNA das mitocôndrias e dos cloroplastos. Os eucariontes apareceram na Terra há aproximadamente 1,5 bilhão de anos.
Ao contrário dos procariontes, que são representados por organismos solitários e formas coloniais, os eucariotos podem ser organismos unicelulares (por exemplo, ameba), coloniais (Volvox) e multicelulares. Eles estão divididos em três grandes reinos: Animais, Plantas e Fungos.
O diâmetro das células eucarióticas é de 5–80 μm. Assim como as células procarióticas, as células eucarióticas são cercadas por membrana de plasma composto por proteínas e lipídios. Esta membrana atua como uma barreira seletiva, permeável a alguns compostos e impermeável a outros. Fora da membrana plasmática existe uma forte parede celular, que nas plantas consiste principalmente em fibras de celulose, e nos fungos - em quitina. A principal função da parede celular é garantir uma forma celular constante. Como a membrana plasmática é permeável à água e as células vegetais e fúngicas normalmente entram em contato com soluções de força iônica inferior à força iônica da solução dentro da célula, a água fluirá para dentro das células. Com isso, o volume das células aumentará, a membrana plasmática começará a se esticar e poderá romper. A parede celular impede a expansão e destruição celular.
Os animais não possuem parede celular, mas a camada externa da membrana plasmática é enriquecida com componentes de carboidratos. Esta camada externa da membrana plasmática das células animais é chamada glicocálice. As células de animais multicelulares não necessitam de uma parede celular forte, uma vez que existem outros mecanismos que proporcionam a regulação do volume celular. Uma vez que as células de animais multicelulares e organismos unicelulares que vivem no mar estão num ambiente em que a concentração total de iões está próxima da concentração de iões intracelulares, as células não incham nem rebentam. Animais unicelulares que vivem em água fresca(ameba, ciliados chinelo), possuem vacúolos contráteis que removem constantemente a água que entra na célula.
Dentro da célula, sob a membrana plasmática, existem citoplasma. A principal substância do citoplasma (hialoplasma) é uma solução concentrada de compostos inorgânicos e orgânicos, cujos principais componentes são as proteínas. Este é um sistema coloidal que pode mudar do estado líquido para o estado gel e vice-versa. Uma parte significativa das proteínas citoplasmáticas são enzimas que realizam vários reações químicas. Localizado no hialoplasma organoides, desempenhando diversas funções na célula. As organelas podem ser membranares (núcleo, aparelho de Golgi, retículo endoplasmático, lisossomos, mitocôndrias, cloroplastos) e não membranares (centro celular, ribossomos, citoesqueleto).
Organelas de membranao principal componente das organelas de membrana é membrana. As membranas biológicas são construídas de acordo com princípio geral, Mas composição química as membranas de diferentes organelas são diferentes. Todas as membranas celulares são filmes finos (7–10 nm de espessura), cuja base é uma dupla camada de lipídios (bicamada), disposta de modo que as partes hidrofílicas carregadas das moléculas estejam em contato com o meio, e os resíduos hidrofóbicos ácidos graxos de cada monocamada são direcionados para dentro da membrana e estão em contato entre si (Fig. 3). As moléculas de proteína (proteínas integrais da membrana) são construídas na bicamada lipídica de tal forma que as partes hidrofóbicas da molécula de proteína ficam em contato com os resíduos de ácidos graxos das moléculas lipídicas e as partes hidrofílicas ficam expostas ao meio ambiente. Além disso, parte das proteínas solúveis (proteínas não membranares) se conecta à membrana principalmente devido a interações iônicas (proteínas de membrana periférica). Fragmentos de carboidratos também estão ligados a muitas proteínas e lipídios nas membranas. Assim, as membranas biológicas são filmes lipídicos nos quais estão incorporadas proteínas integrais.
Uma das principais funções das membranas é criar uma fronteira entre a célula e o ambiente e os vários compartimentos da célula. A bicamada lipídica é permeável principalmente a compostos e gases solúveis em gordura; substâncias hidrofílicas são transportadas através de membranas usando mecanismos especiais: baixo peso molecular - através de vários transportadores (canais, bombas, etc.), e alto peso molecular - através de processos exo- E endocitose(Fig. 4).
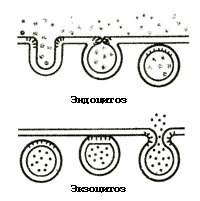
No endocitose certas substâncias são sorvidas na superfície da membrana (devido à interação com proteínas da membrana). Neste ponto, forma-se uma invaginação da membrana no citoplasma. Um frasco contendo o composto transferido é então separado da membrana. Por isso, endocitose- esta é a transferência para a célula de compostos de alto peso molecular do ambiente externo, circundados por uma seção da membrana. O processo inverso, ou seja exocitose- Esta é a transferência de substâncias da célula para o exterior. Ocorre por fusão com a membrana plasmática de uma vesícula cheia de compostos de alto peso molecular transportados. A membrana da vesícula se funde com a membrana plasmática e seu conteúdo é derramado.
Canais, bombas e outros transportadores são moléculas de proteínas de membrana, geralmente formando um poro na membrana.
Além das funções de separar o espaço e garantir a permeabilidade seletiva, as membranas são capazes de detectar sinais. Esta função é desempenhada por proteínas receptoras que se ligam a moléculas sinalizadoras. Proteínas de membrana individuais são enzimas que realizam reações químicas específicas.
Essencial - uma organela celular grande, cercada por um envelope nuclear e geralmente de formato esférico. Existe apenas um núcleo em uma célula e, embora existam células multinucleadas (células músculos esqueléticos, alguns fungos) ou sem núcleo (eritrócitos e plaquetas de mamíferos), mas essas células surgem de células precursoras mononucleares.
A principal função do kernel é armazenamento, transferência e venda de informação genética. Aqui, as moléculas de DNA são duplicadas, fazendo com que, ao se dividirem, as células-filhas recebam o mesmo material genético. No núcleo, utilizando seções individuais de moléculas de DNA (genes) como matriz, ocorre a síntese de moléculas de RNA: informativas (mRNA), de transporte (tRNA) e ribossômicas (rRNA), necessárias à síntese protéica. No núcleo, as subunidades ribossômicas são montadas a partir de moléculas e proteínas de rRNA, que são sintetizadas no citoplasma e transferidas para o núcleo.
O núcleo consiste no envelope nuclear, cromatina (cromossomos), nucléolo e nucleoplasma (carioplasma).

Sob um microscópio, zonas de matéria densa são visíveis dentro do núcleo - cromatina. Nas células que não se dividem, preenche uniformemente o volume do núcleo ou condensa em determinados locais na forma de áreas mais densas e é bem corado com corantes básicos. A cromatina é um complexo de DNA e proteínas (Fig. 5), em sua maioria com carga positiva histonas.
O número de moléculas de DNA no núcleo é igual ao número de cromossomos. O número e a forma dos cromossomos são característica única tipo. Cada cromossomo contém uma molécula de DNA, composta por duas fitas interconectadas e em forma de dupla hélice com 2 nm de espessura. Seu comprimento excede significativamente o diâmetro da célula: pode atingir vários centímetros. A molécula de DNA tem carga negativa, por isso pode dobrar-se (condensar) somente após se ligar a proteínas histonas carregadas positivamente (Fig. 6).
Primeiro, uma fita dupla de DNA gira em torno de blocos de histonas individuais, cada um contendo 8 moléculas de proteína, formando uma estrutura em forma de “contas em um cordão” com espessura de cerca de 10 nm. As contas são chamadas de nucleossomos. Como resultado da formação dos nucleossomos, o comprimento da molécula de DNA diminui aproximadamente 7 vezes. Em seguida, o fio com os nucleossomos se dobra, formando uma estrutura semelhante a uma corda com cerca de 30 nm de espessura. Essa corda, dobrada em voltas, é então presa às proteínas que formam a base do cromossomo. Como resultado, forma-se uma estrutura com espessura de cerca de 300 nm. A condensação adicional desta estrutura leva à formação de um cromossomo.
Durante o período entre as divisões, o cromossomo se desdobra parcialmente. Como resultado disso, seções individuais da molécula de DNA que deveriam ser expressas em uma determinada célula são liberadas de proteínas e esticadas, o que possibilita a leitura de informações delas por meio da síntese de moléculas de RNA.
O nucléolo é um tipo de DNA modelo responsável pela síntese de rRNA e coletado em regiões separadas do núcleo. O nucléolo é a estrutura mais densa do núcleo; não é uma organela separada, mas representa um dos loci do cromossomo. Produz rRNA, que então forma um complexo com proteínas, formando subunidades ribossômicas que vão para o citoplasma.
As proteínas nucleares não-histonas formam uma rede estrutural dentro do núcleo. É representado por uma camada de fibrilas subjacente ao envelope nuclear. Uma rede de fibrilas intranucleares está ligada a ele, à qual estão ligadas fibrilas de cromatina.
O envelope nuclear consiste em duas membranas: externa e interna, separadas por um espaço intermembranar. A membrana externa está em contato com o citoplasma, pode conter polirribossomos e ela própria pode passar para as membranas do retículo endoplasmático. A membrana interna está associada à cromatina. Assim, o envelope nuclear garante a fixação do material cromossômico no espaço tridimensional do núcleo.
O núcleo tem furos redondos - poros nucleares(Fig. 7). Na área dos poros, as membranas externa e interna se fecham e formam buracos preenchidos com fibrilas e grânulos. Dentro do poro está localizado um sistema complexo de proteínas que fornecem ligação seletiva e transferência de macromoléculas. O número de poros nucleares depende da intensidade do metabolismo celular.
Retículo endoplasmático, ou retículo endoplasmático (ER), é uma bizarra rede de canais, vacúolos, sacos achatados, interligados e separados do hialoplasma por uma membrana (Fig. 8).
Distinguir duro E suave RPE . Nas membranas do RE rugoso existem ribossomos(Fig. 9), que sintetizam proteínas excretadas da célula ou integradas na membrana plasmática. A proteína recém-sintetizada sai do ribossomo e passa por um canal especial até a cavidade do retículo endoplasmático, onde sofre modificação pós-tradução, por exemplo, ligação a carboidratos, clivagem proteolítica de parte da cadeia polipeptídica e formação de S –Ligações S entre resíduos de cisteína na cadeia. Essas proteínas são então transportadas para o complexo de Golgi, onde fazem parte dos lisossomos ou dos grânulos secretores. Em ambos os casos, essas proteínas vão parar dentro de uma vesícula membranosa (vesícula).

O RE liso não possui ribossomos. Sua principal função é a síntese lipídica e o metabolismo de carboidratos. Está bem desenvolvido, por exemplo, nas células do córtex adrenal, que contêm enzimas que garantem a síntese dos hormônios esteróides. No RE liso das células do fígado existem enzimas que realizam a oxidação (desintoxicação) de compostos hidrofóbicos estranhos ao organismo, como os medicamentos.

complexo de Golgi (Fig. 10) consiste em 5–10 cavidades planas delimitadas por membrana localizadas em paralelo. As partes finais dessas estruturas em forma de disco possuem extensões. Pode haver várias dessas formações em uma célula. A área do complexo de Golgi contém um grande número de vesículas de membrana. Alguns deles se desprendem das partes terminais da estrutura principal na forma de grânulos secretores e lisossomos. Algumas das pequenas vesículas (vesículas) que transportam proteínas sintetizadas no RE rugoso movem-se para o complexo de Golgi e fundem-se com ele. Assim, o complexo de Golgi está envolvido no acúmulo e posterior modificação de produtos sintetizados no RE bruto e na sua classificação.
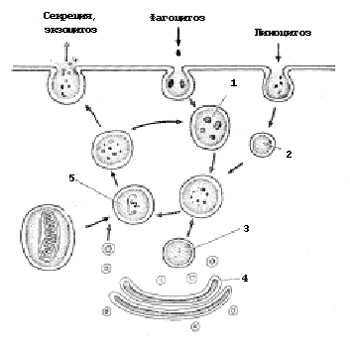
Arroz. 11. Formação e funções dos lisossomas: 1 – fagossoma; 2 – vesícula pinocitótica; 3 – lisossoma primário; 4 – Aparelho de Golgi; 5 – lisossoma secundário
Lisossomos - são vacúolos (Fig. 11), limitados por uma membrana, que brotam do complexo de Golgi. Dentro dos lisossomos o ambiente é bastante ácido (pH 4,9–5,2). Existem enzimas hidrolíticas que decompõem vários polímeros em pH ácido (proteases, nucleases, glicosidases, fosfatases, lipases). Esses lisossomos primários se fundem com vacúolos endocíticos contendo componentes a serem degradados. As substâncias que entram no lisossomo secundário são decompostas em monômeros e transferidas através da membrana do lisossomo para o hialoplasma. Assim, os lisossomos participam dos processos de digestão intracelular.
Mitocôndria rodeado por duas membranas: a externa, que separa a mitocôndria do hialoplasma, e a interna, delimitando seu conteúdo interno. Entre eles existe um espaço intermembranar com 10–20 nm de largura. A membrana interna forma numerosas projeções ( cristas). Esta membrana contém enzimas que garantem a oxidação de aminoácidos, açúcares, glicerol e ácidos graxos formados fora das mitocôndrias (ciclo de Krebs) e realizam a transferência de elétrons na cadeia respiratória (diagrama). Devido à transferência de elétrons ao longo da cadeia respiratória de um nível de energia superior para um inferior, parte da energia livre liberada é armazenada na forma de ATP, a moeda energética universal da célula. Assim, a principal função das mitocôndrias é a oxidação de vários substratos e a síntese de moléculas de ATP.
Dentro da mitocôndria existe uma molécula circular de DNA que codifica parte das proteínas mitocondriais. No espaço interno das mitocôndrias (matriz) existem ribossomos semelhantes aos ribossomos dos procariontes, que garantem a síntese dessas proteínas.
O fato de as mitocôndrias terem seu próprio DNA circular e ribossomos procarióticos levou à hipótese de que a mitocôndria é descendente de uma antiga célula procariótica que uma vez entrou em uma célula eucariótica e, no processo de evolução, assumiu certas funções.
Plastídeos – organelas célula vegetal, que contém pigmentos. EM cloroplastos contém clorofila e carotenóides, cromoplastos– carotenóides, em leucoplastos sem pigmentos. Os plastídios são circundados por uma membrana dupla. Dentro deles existe um sistema de membranas em forma de bolhas planas chamadas tilacóides(Fig. 12). Os tilacóides estão dispostos em pilhas que lembram pilhas de pratos. Os pigmentos estão incorporados nas membranas dos tilacóides. Sua principal função é a absorção de luz, cuja energia, com a ajuda de enzimas incorporadas à membrana do tilacóide, é convertida em um gradiente de íons H + na membrana do tilacóide. Assim como as mitocôndrias, os plastídios possuem seu próprio DNA circular e ribossomos do tipo procariótico. Aparentemente, os plastídios também são organismos procarióticos que vivem em simbiose com células eucarióticas.
Ribossomos são organelas celulares não membranosas encontradas em células pró e eucarióticas. Os ribossomos eucarióticos são maiores em tamanho que os procarióticos, seu tamanho é 25x20x20 nm. O ribossomo consiste em subunidades grandes e pequenas adjacentes umas às outras. Entre as subunidades de um ribossomo funcional existe uma fita de mRNA.
Cada subunidade ribossômica é composta de rRNA, compactado e associado a proteínas. Os ribossomos podem estar localizados livremente no citoplasma ou associados às membranas do RE. Os ribossomos livres podem ser únicos, mas podem formar polissomos quando vários ribossomos estão localizados sequencialmente em uma fita de mRNA. A principal função dos ribossomos é a síntese de proteínas.
Citoesqueleto - Esse sistema musculo-esquelético células, incluindo formações proteicas filamentosas (fibrilares), que são a estrutura da célula e desempenham uma função motora. As estruturas do citoesqueleto são dinâmicas, surgem e se desintegram. O citoesqueleto é representado por três tipos de formações: filamentos intermediários(fios com diâmetro de 10 nm), microfilamento s (fios com diâmetro de 5–7 nm) e microtúbulos. Os filamentos intermediários são estruturas proteicas não ramificadas em forma de fios, muitas vezes dispostos em feixes. Sua composição proteica varia tecidos diferentes: no epitélio consistem em queratina, nos fibroblastos - na vimentina, nas células musculares - na desmina. Os filamentos intermediários desempenharão uma função de estrutura de suporte.
Microfilamentos - são estruturas fibrilares localizadas diretamente sob a membrana plasmática na forma de feixes ou camadas. Eles são claramente visíveis nos pseudópodes da ameba, nos processos móveis dos fibroblastos e nas microvilosidades do epitélio intestinal (Fig. 13). Os microfilamentos são construídos a partir das proteínas contráteis actina e miosina e constituem o aparelho contrátil intracelular.
Microtúbulos fazem parte de estruturas celulares temporárias e permanentes. Os temporários incluem o fuso de divisão, elementos do citoesqueleto das células entre as divisões, e os permanentes incluem cílios, flagelos e centríolos do centro celular. Os microtúbulos são cilindros retos e ocos com um diâmetro de cerca de 24 nm; suas paredes são formadas por moléculas redondas da proteína tubulina. Ao microscópio eletrônico, pode-se observar que a seção transversal dos microtúbulos é formada por 13 subunidades conectadas em um anel. Os microtúbulos estão presentes no hialoplasma de todas as células eucarióticas. Uma das funções dos microtúbulos é criar uma estrutura dentro das células. Além disso, pequenas vesículas se movem ao longo dos microtúbulos, como nos trilhos.
Centro celular consiste em dois centríolos localizados em ângulos retos entre si e microtúbulos associados. Essas organelas nas células em divisão participam da formação do fuso de divisão. A base do centríolo são 9 trigêmeos de microtúbulos localizados ao redor da circunferência, formando um cilindro oco, com 0,2 µm de largura e 0,3–0,5 µm de comprimento. À medida que as células se preparam para se dividir, os centríolos divergem e duplicam. Antes da mitose, os centríolos participam da formação dos microtúbulos fusiformes. As células das plantas superiores não possuem centríolos, mas possuem um centro organizador de microtúbulos semelhante.
A característica mais importante e fundamental das células eucarióticas está associada à localização do aparelho genético na célula. O aparato genético de todos os eucariotos está localizado no núcleo e é protegido pelo envelope nuclear (em grego, “eucarioto” significa ter núcleo). O DNA dos eucariotos é linear (nos procariontes, o DNA é circular e está localizado em uma região especial da célula - o nucleóide, que não é separado por uma membrana do resto do citoplasma). Está associado a proteínas histonas e outras proteínas cromossômicas que as bactérias não possuem.
No ciclo de vida dos eucariotos, geralmente existem duas fases nucleares (haplofase e diplofase). A primeira fase é caracterizada por um conjunto haplóide (único) de cromossomos, então, fundindo-se, duas células haplóides (ou dois núcleos) formam uma célula diplóide (núcleo) contendo um conjunto duplo (diplóide) de cromossomos. Às vezes, durante a próxima divisão, e mais frequentemente após várias divisões, a célula torna-se novamente haplóide. Tal ciclo de vida e, em geral, a diploide não são típicos dos procariontes.
A terceira, talvez a mais interessante diferença, é a presença nas células eucarióticas de organelas especiais que possuem seu próprio aparato genético, se reproduzem por divisão e são circundadas por uma membrana. Essas organelas são mitocôndrias e plastídios. Em sua estrutura e atividade vital, eles são surpreendentemente semelhantes às bactérias. Esta circunstância levou os cientistas modernos a acreditar que tais organismos são descendentes de bactérias que mantiveram uma relação simbiótica com os eucariotos. Os procariontes são caracterizados por um pequeno número de organelas e nenhuma delas é cercada por uma membrana dupla. As células procarióticas não possuem retículo endoplasmático, aparelho de Golgi ou lisossomos.
Outra diferença importante entre procariontes e eucariotos é a presença de endocitose em eucariotos, incluindo fagocitose em muitos grupos. A fagocitose (literalmente “comer por uma célula”) é a capacidade das células eucarióticas de capturar, encerrar em uma vesícula de membrana e digerir uma ampla variedade de partículas sólidas. Este processo fornece uma importante função protetora no corpo. Foi descoberto pela primeira vez por I. I. Mechnikov em uma estrela do mar. O aparecimento de fagocitose em eucariotos está provavelmente associado ao tamanho médio (mais sobre diferenças de tamanho está escrito abaixo). Os tamanhos das células procarióticas são desproporcionalmente menores e, portanto, no processo de desenvolvimento evolutivo dos eucariotos, elas tiveram o problema de fornecer ao corpo uma grande quantidade de alimentos. Como resultado, os primeiros predadores reais e móveis aparecem entre os eucariotos.
A maioria das bactérias possui uma parede celular diferente da eucariótica (nem todos os eucariotos a possuem). Nos procariontes, é uma estrutura durável que consiste principalmente em mureína (em archaea, pseudomureína). A estrutura da mureína é tal que cada célula é cercada por um saco de malha especial, que é uma molécula enorme. Entre os eucariotos, muitos protistas, fungos e plantas possuem parede celular. Nos fungos consiste em quitina e glucanos, nas plantas inferiores consiste em celulose e glicoproteínas, nas diatomáceas sintetizam uma parede celular a partir de ácidos silícicos, nas plantas superiores consiste em celulose, hemicelulose e pectina. Aparentemente, para células eucarióticas maiores tornou-se impossível criar uma parede celular de alta resistência a partir de uma única molécula. Esta circunstância poderia forçar os eucariontes a usar materiais diferentes para a parede celular. Outra explicação é que o ancestral comum dos eucariotos perdeu sua parede celular devido à transição para a predação, e então os genes responsáveis pela síntese da mureína também foram perdidos. Quando alguns eucariontes retornaram à nutrição osmotrófica, a parede celular apareceu novamente, mas em uma base bioquímica diferente.
O metabolismo das bactérias também é diverso. Em geral, existem quatro tipos de nutrição e todos são encontrados entre as bactérias. Estes são fotoautotróficos, fotoheterotróficos, quimioautotróficos, quimioheterotróficos (os fototróficos usam a energia da luz solar, os quimiotróficos usam a energia química).

O que seria do Ano Novo sem champanhe, tangerinas, Olivier, formol e o “arenque com casaco de pele” preferido de todos. Com o último...

Vamos preparar os ingredientes necessários para os biscoitos. A primeira coisa a fazer é colocar a água para ferver. Nós precisamos...

É possível cadastrar funcionário para o cargo de diretor financeiro - contador-chefe? O contador-chefe afirma que sim, mas...

O chefe de uma pequena empresa pode gerenciar facilmente o orçamento de forma independente. VERIFICADO! Se você está fazendo um orçamento...

A criação de novos projetos envolve uma justificação económica preliminar para a sua viabilidade, posterior...
O reporte é gerado pelo RM, é acordado (aprovado) pelo Comité de Risco no âmbito do Conselho de Administração e transmitido a...

Na beira de um prado grande, muito grande, em uma longa folha de grama esmeralda vivia uma pequena joaninha. O pequeno de Deus...

Hoje em dia é bastante comum as pessoas se voltarem para as estrelas. Com a ajuda de um horóscopo uma pessoa pode descobrir...

Negócios ou amigável. Se você estava perseguindo um estranho, isso significa o seu nível de confiança no sexo feminino...

de acordo com o livro dos sonhos de Freud Se você sonhou que estava pescando, isso significa que na vida real dificilmente você consegue desligar...

O turbilhão do Ano Novo nos girará tão rápida e rapidamente que é hora de pensar cuidadosamente sobre o Ano Novo...

O artigo oferece algumas das mais deliciosas receitas para fazer vinagrete. Vinagrete é uma salada deliciosa...

4 de julho de 2015, 20h33 Ultimamente nossa Nastena tem desistido completamente dos doces (e graças a Deus), mas...

Bolo de chocolate perfumado e muito saboroso, é impossível parar de comer! O bolo Beijo do Negro é muito fácil de preparar,...

Vamos preparar os ingredientes necessários para os biscoitos. A primeira coisa a fazer é colocar a água para ferver. Nós...

É possível cadastrar funcionário para o cargo de diretor financeiro - contador-chefe? O contador-chefe afirma...