Шпанська Мушка для двох – як впливають на лібідо у жінок та чоловіків
Біологічно активна добавка на основі екстракту, отриманого з жука шпанкою мушки (або шпанки...)
Щитовидна залоза [glandula thyroidea(PNA), glandula thyreoidea(JNA, BNA)] - непарна залоза внутрішньої секреції. Щитовидна залоза розташовується у передній області шиї; вона синтезує та виділяє в кров та лімфу гормони, що регулюють процеси росту, розвитку, диференціювання тканин та обмін речовин в організмі.
Короткий опис зовнішнього вигляду щитовидної залози вперше дав Гален. Дещо повніше орган описаний у працях А. Везалія (1543). В 1656 Т. Вартон назвав цей орган «щитовидна залоза». У 1836 році Кінг (Th. W. King) першим висунув концепцію про внутрішньосекреторну активність щитовидної залози. Бауманн (Е. Baumann) в 1896 відзначив тісний зв'язок між надходженням йоду в організм і функціональною активністю органу.
Щитовидній залозі вищих хребетних відповідає поджаберний жолобок ланцетника, який йде вентрально по середній лінії вздовж усієї зябрової частини кишечника. У круглоротих щитовидна залоза представлена скупченням одиночних фолікулів, розташованих уздовж краніального відділу кишки. Щитовидна залоза селахій - непарний орган різної формиу амфібій щитовидна залоза парна. У рептилій щитовидна залоза майже завжди непарна, розташовується по серединній лінії, поблизу виходу із серця великих судин і, зазвичай, немає певної форми. У птахів цей орган завжди парний. У ссавців щитовидна залоза розташовується вентрально від каудального відділу гортані та суміжної частини трахеї, складається з двох часток, з'єднаних перешийком у більшості представників даного класу.
Зачаток щитовидної залози виникає у зародка людини на 4-му тижні внутрішньоутробного розвитку (довжина зародка 2,5 мм) у вигляді випинання вентральної стінки глотки по середній лінії між I і II парами зябрових кишень. Це випинання є епітеліальний тяж, який росте вниз вздовж глоткової кишки до рівня III - IV пари зябрових кишень. Епітеліальний тяж на початку свого розвитку є щитомовною протокою (cinctus thyroglossus) і відповідає вивідному протоку щитовидної залози. Потім дистальний конецепітеліальний тяж роздвоюється, і з нього надалі розвиваються права і ліва частки щитовидної залози. Проксимальний кінець епітеліального тяжу атрофується, на його місці в подальшому залишається рудиментарний залишок - сліпий отвір язика (foramen caecum linguae), що локалізується на межі тіла та кореня язика. Таким чином, щитовидна залоза закладається як типова екзокринна залоза, а в процесі подальшого розвиткустає ендокринною (див. Залізи внутрішньої секреції).
Зачатки правої та лівої часткою щитовидної залози, спочатку компактні, швидко збільшуються в обсязі за рахунок розростання епітеліальних клітинних тяжів або трабекул. Між трабекулами проростає мезенхіма із численними кровоносними судинами. На 8-9-му тижні внутрішньоутробного розвитку починають формуватися фолікули, основну масу яких складають тироцити (фолікулярні клітини, А-клітини). Значно менше у складі фолікулів закладається В-клітин (клітин Асканазі). Тироцити та В-клітини близькі між собою. Існує думка, що ці клітини мають спільні стовбурові елементи або можуть трансформуватися одна в одну. У процесі розвитку в зачаток щитовидної залози вростають похідні V пари зябрових кишень - ультимобранхіальні тільця, які є джерелом парафолікулярних клітин, що входять до складу паренхіми щитовидної залози (навколофолікулярних, або С-клітин).
Щитовидна залоза починає функціонувати у плода, що має довжину 7 см, про що свідчить здатність залози, що виникає в цей період, поглинати радіоактивний йод, а також поява колоїду в просвітах фолікулів. Функціонування залози спричиняє диференціювання трабекул, які починають розділятися на окремі невеликі фолікули, що швидко збільшуються в об'ємі в міру накопичення в них колоїду.
Вага (маса) щитовидної залози новонароджених становить середньому 1-2 р. У залозі новонароджених відзначається десквамація фолікулярного епітелію та посилена резорбція колоїду, що, ймовірно, пов'язане з функціональною напругою щитовидної залози в період адаптації до умов навколишнього середовища.
Щитовидна залоза розташовується в передній області шиї (див.) спереду та з боків від трахеї (див.). Вона має підковоподібну форму з увігнутістю, зверненою дозаду, і складається з двох неоднакових за величиною часток (рис. 1). Права (lobus dext.) та ліва (lobus sin.) частки щитовидної залози з'єднані непарним перешийком (isthmus glandulae thyroideae). У тих випадках, коли перешийок відсутня, обидві частки щитовидної залози нещільно прилягають одна до одної.
Іноді зустрічаються додаткові (аберрують) щитовидні залози (glandulae thyroideae accessoriae), або не пов'язані з нею, або з'єднані з частками щитовидної залози невеликими тонкими тяжами. У 30-50% випадків з перешийком або з лівою часткою щитовидна залоза пов'язана пірамідальна частка (lobus pyramidalis), яка, прямуючи вгору, може доходити до верхньої щитовидної вирізки щитовидного хряща або тіла під'язикової кістки (рис. 1).
Вага (маса) щитовидної залози дорослої людини становить 20-60 г. Поздовжній розмір кожної частки досягає 5-8 см, поперечний - 2-4 см, товщина - 1-2,5 см. У період статевого дозрівання щитовидна залоза збільшується. Розміри її можуть змінюватись також залежно від ступеня кровонаповнення; в старечому віцірозміри щитовидної залози зменшуються.
Зовні щитовидна залоза покрита фіброзною капсулою, яка пов'язана сполучнотканинними пучками з перстнеподібним хрящем, кільцями трахеї. Найбільш щільні сполучнотканинні пучки утворюють своєрідні зв'язки. Серед них особливо виражені середня зв'язка щитовидної залози, яка тягнеться від капсули перешийка до передньої поверхні перстневидного хряща, а також права і ліва латеральні зв'язки щитовидної залози, що розташовуються між капсулою нижньомедіальних ділянок бічних часток, латеральними поверхнями перстневид еі.

Переднебоковые поверхности щитовидной железы покрыты грудиноподъязычными (mm. sternohyoidei) и грудинощитовидными мышцами (mm. sternothyroidei), верхними брюшками правой и левой лопаточно-подъязычных мышц (mm. omohyoidei dext. et sin.), лежащих между листками предтрахеальной пластинки шейной фасции. На межі передньо-бічної та задньомедіальної поверхонь щитовидної залози до неї прилягає судинно-нервовий пучокшиї (цветн. рис. 3). По задньомедіальній поверхні щитовидної залози проходить зворотний гортанний нерв (n. laryngeus recurrens) і розташовуються паратрахеальні лімфатичні вузли. Заднемедіальні поверхні залози прилягають до бокових поверхонь верхніх кілець трахеї, глотки і стравоходу, а вгорі - до перстневидного і щитовидного хрящів.
Кровопостачання здійснюється з верхніх щитовидних артерій (аа. thyroideae sup. dext. etsi п.), що відходять від зовнішніх сонних артерій (аа. carotides ext.) і з нижніх щитовидних артерій (аа. thyroideae inf. dext. від щитошийних стовбурів (trunci thyrocervicales). Приблизно в 10% випадків у кровопостачанні щитовидної залози бере участь нижча щитовидна артерія (a. thyroidea ima), що відходить від плечеголовного стовбура (truncus brachiocephalicus) або від дуги аорти (arcus aortae), рідше - від загальної. На поверхні залози артерії утворюють мережу анастомозирующую (цветн. рис. 4,5), яка розпадається на капіляри, що оточують фолікули і тісно прилеглі до фолікулярного епітелію. Венозна кроввідтікає через однойменні вени у внутрішню яремну вену (v. jugularis interna) та плечеголовні вени (vv. brachiocephalicae).
Лімфовідтікання відбувається через лімфатичні судини, що впадають у паратрахеальні, глибокі шийні та середостінні лімфатичні вузли. Лімфатичні капіляри і невеликі лімфатичні судини лежать Е1 безпосередньо між фолікулами.
Іннервація. Симпатична іннервація щитовидної залози здійснюється нервами, що йдуть від шийних вузлів симпатичних стовбурів. Парасимпатична іннервація забезпечується гілками блукаючого нерва (див.) – верхнім гортанним (n. laryngeus sup.) та зворотним гортанним (n. laryngeus recurrens) нервами.
Від фіброзної капсули, що покриває щитовидну залозу, в глиб залози відходять сполучнотканинні перегородки, які утворюють строму залози і містять у своїй товщі судини та нерви. Ці сполучнотканинні перегородки не з'єднуються один з одним у глибині тканини щитовидної залози. Тому поділ паренхіми на часточки неповний, і заліза є псевдодольчастою. Щитовидна залоза має гістологічну будову, типову для залоз внутрішньої секреції: у ній відсутні вивідні протоки і кожна функціональна одиниця тісно пов'язана з кровоносною системою. Структурною одиницею щитовидної залози є фолікул - округлий або злегка овальний замкнутий пляшечку, стінка якого вистелена секреторним (фолікулярним) епітелієм.
У паренхімі щитовидної залози розрізняють три види клітин (А, В та С), які відрізняються один від одного як у структурному, так і у функціональному відношенні. Основну масу клітин паренхіми щитовидної залози становлять тироцити (фолікулярні клітини, або А-клітини), що виробляють тиреоїдні гормони. Залежно від функціонального стану щитовидної залози, тироцити можуть бути плоскими, кубічними або циліндричними. При низькій функціональній активності щитовидної залози тироцити, як правило, плоскі, при високій – циліндричні.
Просвіт фолікула заповнений колоїдом, який являє собою гомогенну масу, що забарвлюється гематоксилін-еозином рожевий колір. За даними електронної мікроскопії (див.), колоїд має дрібнозернисту структуру та середню електронну щільність. Основну масу колоїду становить тиреоглобулін (див.), що секретується тироцитами, характерною особливістю яких є активне захоплення йоду (див.). Колоїд безпосередньо прилягає до апікальної поверхні тироцитів (апікальної мембрани), на якій є численні мікроворсинки. Розташовані поруч тироцити з'єднані один з одним за допомогою замикаючих пластин, або термінальних перемичок, і десмосом. На базальній поверхні тироцитів можуть з'являтись глибокі складки, особливо виражені в період функціональної активності, які значно збільшують поверхню клітин, звернену до кровоносних капілярів. Між тироцитами та кровоносними капілярамизнаходяться базальна мембрана, основна речовина, тонкі колагенові та ретикулярні волокна, орієнтовані у різних напрямках.

У цитоплазмі тироцитів добре розвинена зерниста ендоплазматична мережа (див. Ендоплазматичний ретикулум). Мітохондрії розташовані по всій клітині, проте в апікальній частині їх завжди дещо більше, ніж у базальній частині клітини. Простежується чіткий топографічний зв'язок мітохондрій із трубочками зернистої ендоплазматичної мережі. Так, останні нерідко «огортають» окремі мітохондрії. У той же час мітохондрії можуть частково або повністю охоплювати окремі елементи ендоплазматичної мережі. У тироцитах добре розвинений комплекс Гольджі (див. Гольджі комплекс), який представлений великими вакуолями, сплощеними цистернами (вакуолями) та мікробульбашками. Усередині кільця комплексу Гольджі, а також поблизу нього виявляються гранули різної величини і форми, різної електронної щільності, що виявляються при введенні радіоактивного йоду (рис. 2, а). Подібні гранули є не лише поблизу комплексу Гольджі, але й інших ділянках клітини; наприклад, в її апікальній частині вони іноді утворюють цілі скупчення, що складаються з кількох рядів гранул (від 3 до 8), розташованих один під одним. Крім характерних гранул, в апікальній частині тироцитів іноді виявляються внутрішньоклітинні краплі колоїду.
В-клітини (клітини Асканазі) - більші, ніж тироцити, мають еозинофільну цитоплазму та округле центрально розташоване ядро. Вони містять велику кількість овальних або округлих мітохондрій, серед яких розташовані секреторні гранули. У цитоплазмі цих клітин виявлено біогенні аміни, зокрема серотонін (див.). Вперше В-клітини з'являються у віці 14-16 років. У великій кількості вони зустрічаються у людей віком 50-60 років.
Парафолікулярні клітини (навколофолікулярні, або С-клітини, або К-клітини) відрізняються від тироцитон відсутністю здатності поглинати йод. Вони забезпечують синтез кальцитоніну (див.) – гормону, що бере участь в обміні кальцію в організмі. Окремі парафолікулярні клітини або їх групи локалізуються на зовнішній поверхні фолікулів (рис. 2). Вони ніколи не контактують із колоїдом, від якого відокремлені цитоплазмою тироцитів. Парафолікулярні клітини мають порівняно великі розміри, низьку електронну густину цитоплазми, яка густо заповнена білковими гранулами, що виявляються при срібло (рис. 2, б). У парафолікулярних клітинах добре розвинені зерниста ендоплазматична мережа та комплекс Гольджі.
Поряд з фолікулами в щитовидній залозі розрізняють інтерфолікулярні (екстрафолікулярні) острівці, утворені клітинами, структура яких нагадує будову типових тироцитів. У центрах деяких інтерфолікулярних острівців є мікрофолікули, які з кількох клітин. До складу інтерфолікулярних острівців входять парафолікулярні клітини. Найбільш часто парафолікулярні клітини зустрічаються в острівцях, розташованих у центральній частині залози, де вони становлять близько 2-5% всіх клітин. Інтерфолікулярні острівці мають значення у регенерації тканини щитовидної залози у разі, якщо поразка останньої має великий характер і супроводжується загибеллю цілих фолікулів. При часткових ушкодженнях фолікулів регенерація здійснюється з допомогою тироцитів, розташованих базально. Завдяки останнім відбувається фізіологічна регенерація фолікулярного епітелію.
Фізіологічна роль щитовидної залози полягає в біосинтезі та виділенні в кров та лімфу гормонів, що регулюють процеси росту, розвитку, диференціювання тканин та активують обмін речовин в організмі. Специфічною особливістю тироцитів є здатність активно поглинати, накопичувати йод і переводити його в органічно пов'язану форму шляхом утворення йодовмісних тиреоїдних гормонів - тироксину (див.) та трийодтироніну (див.).
Секреторний процес, що протікає у щитовидній залозі, складається з трьох фаз. У першій фазі (фаза продукції) відбувається утворення тиреоглобуліну, а також окиснення йодидів в атомарний йод. У другій фазі (фаза виділення, або секреції) тиреоглобулін виділяється у просвіт фолікула, конденсується у ньому у вигляді колоїду та йодується. Третя фаза (фаза виведення) складається з реабсорбції колоїду тироцитами, транспортування реабсорбованих речовин через цитоплазму до базальної частини тироциту та вивільнення тиреоїдних гормонів у кров.
Фаза продукції тиреоглобуліну починається з накопичення зернистої ендоплазматичної мережі тироциту вихідних амінокислот, що надходять з крові. Під впливом матричної РНК (див. Рибонуклеїнові кислоти), що міститься в рибосомах, відбувається синтез первинного поліпептиду, що накопичується в лакунах ендоплазматичної мережі. Тут починається приєднання до поліпептиду вуглеводів (галактози і маннози). Синтезований первинний поліпептид переміщається до комплексу Гольджі, де завершується його гліколізування, відбувається складання та упаковка молекул глікопротеїду, що входять до складу тиреоглобуліну (див.). Секреторні бульбашки, що формуються в зоні комплексу Гольджі та містять глікопротеїд (нейодований тиреоглобулін), зміщуються в апікальну частину тироциту, зливаються своїми оболонками з його апікальною мембраною та виділяють вміст у просвіт фолікула шляхом екзоцитозу.
Йод надходить у тироцити з крові у вигляді йодиду (йод-іону), транспортується через їхню цитоплазму і виділяється через апікальну мембрану в просвіт фолікула, заповнений колоїдом.
Поглинання йоду тироцитами розглядається як активний процес перенесення йодиду проти градієнта концентрації, що потребує витрати енергії. Такий високоактивний і високоспецифічний транспорт йоду, а також (гострий переведення цього елемента у зв'язану форму визначають роль щитовидної залози як головного органу обміну йоду в організмі.) Зміст йоду в щитовидній залозі перевищує його рівень в інших тканинах та сироватці крові 10-100 разів.
Оскільки брати участь у процесі йодування тиреоглобуліну може лише атомарний йод, то йодид зазнає окислення, яке здійснюється в субапікальній зоні цитоплазми тироцитів за участю пероксидази (див. Пероксидази).
Фаза виділення, або секреції, тиреоїдних гормонів починається з виходу нейодованого тиреоглобуліну в просвіт фолікула та надходження його в колоїд. Йодування тиреоглобуліну (включення атомів йоду до тирозильних радикалів) відбувається в периферичній зоні фолікулів, на межі апікальної частини тироциту та колоїду. До числа амінокислот, що становлять білковий компонент молекули тиреоглобуліну, входять тирозин (див.) та його похідні - тироніни, які, зазнаючи йодування, дають початок тиреоїдним гормонам: тироксину (Т4) та трийодтироніну (Т3). Детально синтез тиреоїдних гормонів - див.
Поряд із тиреоглобуліном у щитовидній залозі утворюється тиреоальбумін, який теж йодується, але лише частково, до стадії йодтирозинів. У нормі співвідношення концентрацій тиреоглобуліну та тиреоальбуміну становить приблизно 9:1. При патологічних станах, що супроводжуються проліферацією паренхіми щитовидної залози, її зобною трансформацією та виникненням аденом, утворення тиреоальбуміну збільшується, а при злоякісних пухлинах щитовидної залози може навіть перевершувати утворення тиреоглобуліну. Крім того, у щитовидній залозі виявлено йодовані гістидини та тироксамін. Усі йодовані амінокислоти, що входять до складу тиреоїдних білків, є L-ізомерами (див. Ізомерія).
Заключна (третя) фаза секреторного процесу, що протікає в тироцитах, - фаза виведення тиреоїдних гормонів із фолікулів у кров. Оскільки тиреоїдні гормони містяться у молекулах тиреоглобуліну у зв'язаному стані, потреба організму в них може бути задоволена лише шляхом розщеплення молекули тиреоглобуліну. Фаза виведення супроводжується значним посиленням процесів дисиміляції в тироцитах (про що свідчить виразне збільшення поглинання ними кисню) та сильним набуханням їх цитоплазми та ядер. Фаза виведення починається із реабсорбції колоїду тироцитами. Електронно-мікроскопічні дослідження дозволили встановити, що реабсорбція колоїду здійснюється шляхом його активного фагоцитозу тироцитами за допомогою псевдоподій (макроендоцитоз). У цитоплазмі тироцитів з'являються краплі колоїду, до яких наближаються лізосоми та зливаються з ними. Тиреоглобулін у краплях колоїду розщеплюється під дією ферментів лізосом (див.), в результаті чого вивільняються йодтирозини: монойодтирозин і дийодтирозин і йодтироніни (тироксин і трийодтиронін), що накопичуються в зміщуються в базальну частину тироцита. Йодтирозини при цьому повністю дейодуються і в кров не надходять, а йод, що звільняється з них, знову використовується в біосинтезі тиреоїдних гормонів. Йодтироніни після випорожнення вакуолів потрапляють через базальну мембрану та перикапілярний простір у кровоносні (частково і в лімфатичні) капіляри, що обплітають фолікул.
Поява псевдоподій та крапель колоїду у тироцитах спостерігається лише у початковому періоді фази виведення. Надалі при нормальній функції щитовидної залози процеси виділення протікають без посиленого утворення псевдоподій та крапель колоїду шляхом піноцитозу (мікроендоцитозу). Зазначені механізми набувають чинності послідовно: у початковому періоді фази виведення переважає макроендоцитоз псевдоподіями, надалі він змінюється мікроендоцитозом.
Вихід у кров тиреоїдних гормонів із щитовидної залози, наведеної у стан гіперфункції повторними впливами тиреотропного гормону, з самого початку протікає у вигляді мікроендоцитозу без утворення псевдоподій та чітко помітних крапель інтрацелюлярного колоїду. Такі ж співвідношення визначаються при тиреотоксикозі, коли високий рівень тироксину і трийодтироніну в крові свідчить не тільки про збільшення продукції тиреоїдних гормонів, але і про інтенсивне виведення їх у кров; при цьому не виявляється ні псевдоподій, ні чітких крапель інтрацелюлярного колоїду.
Крім йодованих тиреоїдних гормонів, щитовидна залоза утворює кальцитонін (див.) - безйодний білковий гормон, що знижує вміст кальцію в крові. Кальцитонін утворюється парафолікулярними клітинами. Парафолікулярні клітини є за своїм походженням видозміненими нервовими клітинами(нейроендокринними) та зберігають здатність поглинати попередників нейроамінів (L-ДОФА та 5-гідрокситриптофан) та декарбоксилювати їх відповідно у нор-адреналін (див.) та серотонін. Високий вміст нейроамінів та здатність виробляти білковий гормон зумовлюють включення парафолікулярних клітин щитовидної залози в АПУД-систему (див. АПУД-система). Парафолікулярні клітини потенціюють діяльність фолікулярного епітелію та сприяють підтримці внутрішньоорганного гомеостазу щитовидної залози.
Функція В-клітин визначається накопиченням біогенних амінів, зокрема серотоніну, та потенціюванням фізіологічної активності фолікулярного епітелію.
Специфічним стимулятором щитовидної залози вважається тиреотропний гормон гіпофіза. Тиреотропна функція передньої частки гіпофіза, у свою чергу, активується тироліберином, що секретується гіпоталамусом (див. Гіпоталамічні нейрогормони). Тому пошкодження гіпоталамуса призводить до такого ж ослаблення діяльності щитовидної залози, як і гіпофізектомія (див. Гіпофіз). Цей спосіб регуляції може бути позначений як трансаденогіпофізарний.
У свою чергу, тиреоїдні гормони (особливо трийодтиронін) пригнічують тиреотропну функцію гіпофіза (і, імовірно, секрецію тироліберину гіпоталамусом), тобто взаємовідносини між функціональною активністю щитовидної залози і інтенсивністю тиреотропної функції гіпофіза представляють систему . функціональної активності щитовидної залози у межах фізіологічної норми.
Тиреотропний гормон, що надходить до щитовидної залози зі струмом крові, сприймається специфічними рецепторами, що локалізуються у плазматичній мембрані тироцитів. Ці рецептори, з'єднавшись з тиреотропним гормоном, активують аденілатциклазну систему тироцитів, яка за допомогою циклічного аденозинмонофосфату (цАМФ) пускає в хід ферментні системи тироцитів, внаслідок чого збільшується їхня функціональна активність.
Встановлено, що секреція тиреоїдних гормонів активується безпосередньо симпатичними імпульсами, хоча й настільки інтенсивно, як тиреотропним гормоном. Парасимпатичні імпульси викликають гноблення цих процесів. Таким чином, регулюючі впливи гіпоталамуса на щитовидну залозу можуть відбуватися як через гіпофіз, так і минаючи його (парагіпофізарно).
У той же час аферентні сигнали зі щитовидної залози, що надходять по доцентрових нервових шляхах, досягнувши гіпоталамуса, послаблюють тиреотропну функцію гіпофіза; отже, негативна Зворотній зв'язокміж щитовидною залозою та гіпофізом проявляється також і в прямої дії нервових імпульсів. Стан та діяльність парафолікулярних клітин щитовидної залози не залежать від гіпофіза та не порушуються після гіпофізектомії; їхню функцію стимулюють симпатичні імпульси, а парасимпатичні пригнічують. Разом з тим секреторна активність парафолікулярних клітин має пряму залежність від концентрації кальцію в крові: підвищення або зниження її спричиняє відповідно збільшення або ослаблення секреції кальцитоніну парафолікулярними клітинами. Антагоністично взаємодіючи з паратиреоїдним гормоном (див. Паратгормон) паращитовидних залоз (див. Паращитовидні залози), кальцитонін забезпечує збереження сталості рівня кальцію в організмі.
Майже весь тироксин, що надходить у кров, оборотно зв'язується з білками сироватки, переважно з L-глобуліном - так званим тіроксинзв'язуючим глобуліном, а частково з тіроксинзв'язуючим преальбуміном і альбуміном. Тому концентрацію білково-зв'язаного йоду в крові часто розглядають як показник секреторної активності щитовидної залози. Зв'язування тироксину з білками сироватки крові запобігає його руйнуванню, але перешкоджає його активної діїна клітини. Між пов'язаним і вільним тироксином у крові встановлюється динамічна рівновага, і лише вільний тироксин впливає на клітини, що реагують, і тканини. Трийодтиронін зв'язується з білками сироватки слабше, ніж тироксин. Напіврозпад тироксину, що знаходиться в крові, триває 6-7 діб, трийодтиронін розпадається швидше (період напіврозпаду 2 діб).
Рецепція тироксину відбувається усередині клітин. Проникнувши в клітину, тироксин одразу втрачає один атом йоду, переходячи в трийодтиронін. Точкою застосування трийодтироніну (як що надійшов з крові, так і утвореного з тироксину) є ДНК, де трийодтиронін стимулює транскрипцію (див.) та утворення РНК.
У клітинах відбувається подальше дейодування тироксину та трийодтироніну, дезамінування, розрив дифенілзфірного зв'язку та декарбоксилювання (див. Йодний обмін).
У метаболізмі тиреоїдних гормонів головна роль належить печінці, в якій продукти розпаду йодтиронинів, що дейодуються, зв'язуються в глюкуронові і сірчані кон'югати і потім з жовчю надходять в кишечник, звідки звільнився йод всмоктується назад в кров, переноситься до й.
В основі ефектів, що викликаються тиреоїдними гормонами, лежить їхній вплив на поглинання кисню та окислювальні процеси в організмі. Встановлено, що тироксин у токсичних дозах діє на мітохондрії клітин, роз'єднуючи синтез АТФ з перенесенням електронів по дихальному ланцюгу та блокуючи таким чином окислювальне фосфорилювання (див.).
Тиреоїдні гормони посилюють теплоутворення, і за їх недостатності (гіпотиреозі) знижується температура тіла. Одночасно гіпотиреоз (див.) супроводжується затримкою води в організмі та зменшенням виділення кальцію та фосфору із сечею.
Тиреоїдні гормони посилюють розпад глікогену (див.) та зменшують його утворення у печінці. Недостатність цих гормонів супроводжується розладом регуляції вуглеводного обміну (див.) та підвищенням толерантності організму до глюкози. При гіпертиреозі (див. Тиреотоксикоз) збільшується виділення азоту з сечею і порушується (фосфорилювання креатину в умовах гіпотиреозу підвищується вміст холестерину в крові, а при надлишку тиреоїдних гормонів його рівень знижується. У той же час при гіпертиреозі посилюється збудливість вищої нервової системи (особливо її симпатичного відділу), що проявляється тахікардією (див.), аритміями (див. Аритмії серця), збільшенням швидкості кровотоку, підвищенням систолічного артеріального тиску, одночасно посилюється моторика шлунково-кишкового тракту та секреція травних соків.
Тиреоїдні гормони необхідні нормального функціонування центральної нервової системи. Недостатність тиреоїдних гормонів в ембріональному періоді та на початку постнатального періоду може призвести до затримки диференціювання кори головного мозку та психічного розвиткудитини до кретинізму (див.).
Тиреоїдні гормони разом із соматотропним гормоном (див.) беруть участь у регуляції росту тіла (особливо стимулюють окостеніння).
Під час вагітності функціональна активність щитовидної залози матері посилюється; збільшення вмісту у крові загального тироксину пов'язані з посиленням синтезу тиреотропного гормону під впливом плацентарних естрогенів.
Здатність щитовидної залози до концентрації та накопичення йоду з'являється у плода на 10 – 12-му тижні внутрішньоутробного розвитку. У цей час починається синтез монойодтироніну, дийодтироніну, трийодтироніну, тироксину, тироксисвязывающего глобуліну. У сироватці крові плода з'являються тіроліберин (тиреотропін-рилізинг-гормон) та тиреотропний гормон гіпофізарного походження. Регуляторні взаємини між тиреотропним гормоном та тиреоїдними гормонами встановлюються з 30-го тижня внутрішньоутробного розвитку.
Паралелізму між вмістом тиреотропного та тиреоїдних гормонів у крові матері та плоду не виявлено, оскільки трансплацентарний транспорт цих гормонів становить менше 1%. Найвища концентрація тиреоїдних гормонів у внутрішньоутробному періоді виявляється у плода перед його народженням.
Відразу після народження відзначається період підвищеної функціональної активності щитовидної залози. Рівень тиреотропного гормону підвищується на 30 хвилині після народження, а через 24-48 годин знижується до такого ж рівня, як у дорослих. Вміст трийодтироніну максимально збільшується до кінця першої доби. Максимальне збільшення вмісту тироксину відзначається через 24-48 годин після народження, потім відбувається поступове зниження рівня.
У недоношених дітей підвищення вмісту тиреотропного гормону і тиреоїдних гормонів менш виражено, особливо у дітей з низькою вагою тіла при народженні. Проте протягом кількох тижнів після народження таких дітей відбувається зниження рівня тиреоїдних гормонів, як і у доношених. І у доношених, і у недоношених дітей при різних захворюванняхрівень тиреотропного та тиреоїдних гормонів може бути значно знижений, проте протягом кількох тижнів він нормалізується.
Функціональна активність щитовидної залози тривалий час зберігається на стабільному рівні. Лише у старечому віці спостерігаються атрофічні зміни в паренхімі залози, що супроводжуються невеликим зниженням рівня загального обміну, проте відзначаються ознаки посилення функціональної активності щитовидної залози, що можна розглядати як компенсаторну реакцію, що протидіє ослабленню окислювальних процесів у тканинах організму, що старіє.
Дистрофія може спостерігатись при порушеннях тканинного (клітинного) обміну щитовидної залози, головним чином в умовах патології. Такі її види, як зерниста (паренхіматозна) та гідропічна (див. Вакуольна дистрофія) дистрофія тироцитів є різновидом білкової дистрофії (див.). При зернистій дистрофії в цитоплазмі тироцитів з'являються включення білкової природи, відзначається набухання мітохондрій, сплощення їх христ, виявляється розширення цистерн ендоплазматичної мережі, накопичення білка. При гідропічній дистрофії у цитоплазмі тироцитів, рідше в ядрі, з'являються вакуолі, заповнені рідиною.
Амілоїдоз щитовидної залози зустрічається рідко. Він спостерігається при генералізованому амілоїдозі (див.) і характеризується відкладенням амілоїду в стромі залози, базальній мембрані фолікулів, стінках кровоносних та лімфатичних судин. Відкладення амілоїду характерне для медулярного раку щитовидної залози. Доведено участь у освіті амілоїду епітеліальних пухлинних клітин.
Заміщення паренхіми щитовидної залози жировою тканиною спостерігається при атрофії щитовидної залози, особливо при так званій гормональній атрофії, що супроводжується зниженням функції залози, наприклад, при апітуїтаризмі, мікседемі. Описано також повне природжене заміщення щитовидної залози жировою тканиною.
Мінеральні дистрофії щитовидної залози (кальцинози) можуть бути внутрішньоклітинними та позаклітинними, характеризуються випаданням солей кальцію у вигляді зерен різної величини у некротизованих або дистрофічно змінених клітинах та структурах. Матрицею внутрішньоклітинного звапніння є мітохондрії та лізосоми тироцитів, а позаклітинного (найпоширенішого) - колагенові волокна строми. Причиною кальцинозу є місцеві фактори, а також загальні, такі як гіперкальціємія, що виникає при нестачі кальцитоніну, при гіперпродукції паратгормону, посиленому виході кальцію з депо, зменшенні виділення кальцію з організму.
Порушення обміну пігментів у щитовидній залозі, зокрема гемоглобіногенних, спостерігається у вогнищах крововиливів при гемосидерозі (див.) та гемохроматозі (див.). При цьому гемосидерин і феритин виявляються по ходу стромових волокон, в цитоплазмі клітин.
Некроз тканини щитовидної залози у вигляді ішемічного інфаркту розвивається при перев'язці щитовидних артерій або їх тромбозі, при атеросклерозі, новоутвореннях органів шиї. Дрібні некрози щитовидної залози спостерігаються при різних варіантах зоба, при тиреоїдиті, внаслідок порушення кровообігу, при опроміненні.
Порушення кровообігу проявляються розладами кровонаповнення щитовидної залози, тромбозом її судин, емболією, інфарктом. Найчастіше спостерігається колатеральна гіперемія (при утрудненні кровотоку внаслідок гіперплазії тканини щитовидної залози або зростання її пухлини). Тривалий застій крові в щитовидній залозі призводить до загибелі її паренхіми та супроводжується безклітинним склерозом. Наслідком порушень гемодинаміки, що спостерігаються при родовій травмі, артеріальній гіпертензії, системних васкулітах, інфекційних хворобах (тифи, сепсис), лейкозі, анемії, є крововиливи (див.), плазморагія (див.). Плазморагія у щитовидній залозі спостерігається при порушенні проникності судин мікроциркуляторного русла (див. Мікроциркуляція). Мікроскопічно відзначається сплощення ендотелію судин, фібриноїдне набухання (див. Фібриноїдне перетворення) та некроз судинної стінки.
Запалення щитовидної залози спостерігається рідко; може виникати при ангіні, остеомієліті, сепсисі, а також при деяких специфічних інфекційних захворюваннях (наприклад, туберкульозі, сифілісі, актиномікозі). Воно може протікати гостро, підгостро та хронічно. Гострий гнійний тиреоїдит характеризується формуванням у щитовидній залозі дрібних або великих абсцесів. Великі абсцеси можуть розкриватися середостіння, трахею і через шкіру з утворенням свищів. Специфічні тиреоїдити (туберкульозний, сифілітичний, актиномікозний) зустрічаються рідко, як прояв загального захворювання (див. Тиреоїдит).
Кісти різного розміру найчастіше виявляються у зобнозміненій щитовидній залозі; вони виникають як наслідок колишніх крововиливів та стазу колоїду (фолікулярні кісти), а також внаслідок вад розвитку ультимобранхіальних тілець (ультимобранхіальні кісти). Кісти, особливо фолікулярні, вистелені кубічним або плоским епітелієм і мають потовщену фіброзну стінку.
Атрофія щитовидної залози спостерігається в старечому віці, іноді при цукровому діабеті, гіповітамінозі В, гіперплазії надниркових залоз, захворюваннях гіпофіза та ін. Атрофія щитовидної залози характеризується зменшенням її ваги (маси), числа та розмірів фолікулів та клітин. Атрофія паренхіми щитовидної залози може супроводжуватися заміщенням тканини залози сполучною тканиною. Іноді в осередках склерозу відзначається метаплазія (див.) циліндричних тироцитів у плоскі (епідермоїдна метаплазія).
Гіперплазія тканини щитовидної залози в період статевого дозрівання пов'язана із зміною функції статевих залоз. В умовах патології гіперплазія обумовлена надмірною секрецією тиреотропного гормону гіпофіза. Вона може бути дифузною та вогнищевою. При гіперплазії відзначається посилена проліферація клітин інтерфолікулярних острівців з утворенням нових фолікулів та тироцитів, що утворюють сосочкові вирости та так звані сандерсонові подушки (див. Зоб спорадичний). Спостерігається збільшення висоти тироцитів, накопичення в них рибонуклеопротеїдів, йодидпероксидази у навколоядерній зоні, тиреоглобуліну в апікальних відділах клітини. Характерно збільшення розмірів ядер, кількості та розмірів цитоплазматичних органел. Виявляється гіперплазія фібрилярних структур базальної мембрани фолікулів, кровоносних капілярів. У фолікулах може спостерігатися розрідження та посилена резорбція колоїду (при дифузному токсичному зобі).
Методи обстеження хворих із захворюваннями щитовидної залози включають клінічне обстеження та методи оцінки функції та структури щитовидної залози.
Клінічне обстеження є важливою ланкою у діагностиці захворювань щитовидної залози. Воно складається зі збору скарг, анамнезу та об'єктивних даних (стан шкіри, підшкірної клітковини, волосся, нервово-м'язової та серцево-судинної систем, шлунково-кишковий тракт). Особлива увага приділяється пальпації щитовидної залози, яка дає інформацію про розміри, симетрію часток та консистенцію органу.
Функцію щитовидної залози оцінюють за допомогою непрямих та специфічних методів. Непрямі методи засновані на дослідженні фізіологічних функцій організму, на які впливають тиреоїдні гормони. Показники, одержувані з допомогою цих методів, неспецифічні для патології щитовидної залози, оскільки подібні зміни можуть бути і при захворюваннях інших органів. Непрямі методи включають дослідження основного обміну (див. Обмін речовин та енергії), жирового (вміст у крові холестерину та неетерифікованих жирних кислот) та білкового обміну, стану нервово-м'язової (див. Рефлексометрія) та серцево-судинної (див. Електрокардіографія) систем.
Специфічні методи оцінки функціонального стану щитовидної залози включають дослідження рівня тиреоїдних гормонів у крові та обміну йоду (див. Йодний обмін). Для визначення тиреоїдних гормонів використовують різні методи, зокрема і біохімічні. Останні дозволяють встановити концентрацію в крові йоду, пов'язаного білками плазми (див. Білково-пов'язаний йод), і йоду, що екстрагується бутанолом (див. Бутанолекстрагований йод). Хімічні методивизначення тиреоїдних гормонів трудомісткі та складні. З використанням імунологічних методів вони втратили своє значення і використовуються лише у спеціальних лабораторіях.
У основі імунологічних методів лежить принцип конкурентного зв'язування гормонів та інших досліджуваних речовин специфічними антитілами. Як мітки використовують радіонуклід (див. Радіоімунологічний метод). В даний час ці методи застосовують для визначення в сироватці крові загального та вільного тироксину (Т4), загального, вільного та реверсивного, або зворотного, трийодтироніну (Т3), тироксин-зв'язуючого глобуліну (ТСГ), тиреотропного гормону (ТТГ), тироліберину (ТТРГ) ) та тиреоглобулінових антитіл. Дослідження проводять in vitro за допомогою спеціальних тест-наборів за стандартною методикою.
До специфічних методів оцінки обміну йоду належать також радіонуклідні методи з використанням 123 I, 125 I, 131 I, 132 I та 99m Tc-пертехнетату (див. Радіофармацевтичні препарати). Абсолютних протипоказаньдля застосування зазначених радіонуклідів не існує, до відносних відносять дитячий вік, вагітність та період годування дитини грудьми, а при застосуванні радіоактивного йоду знижену функцію щитовидної залози. За 1,5 - 2 місяці до дослідження скасовують усі йодовмісні та бромовмісні препарати, антитиреоїдні, седативні засоби, гормони, введення рентгеноконтрастних сполук йоду, змазування шкіри спиртовим розчиномйоду; з харчового раціону виключають продукти, багаті на йод ( морську капустута рибу, мінеральні води, хурму та ін.). Для вивчення внутрішньотиреоїдного обміну йоду використовують тест накопичення щитовидної залози радіоактивного йоду та 99тТс-пертех-нетата. Для цього хворому дають всередину або внутрішньовенно вводять 0,0025-0,005 мкюрі (0,1-0,2 МБк) 131 I, 125 I або 0,001-0,02 мкюрі (0,4- 0,8 МБк) 123 I, 132 I, або 1 мкюр (40 МБк) 99m Tc-пертехнетату. Гамма-випромінювання реєструють за допомогою одноканальної радіометричної установки, датчик якої мають 25-30 см від передньої поверхні шиї хворого. Інтенсивність випромінювання над щитовидною залозою реєструють через 2,4 та 24 години після прийому або введення радіонукліду. Отримані результати радіометрії порівнюють із загальною активністю введеного в організм радіонукліду, що приймається за 100%. У здорових осіб показники накопичення щитовидної залози радіоактивного йоду через 2 години не перевищують 20%, через 24 години - 50%, накопичення 99m Tc-пертехнетату через 2 години не перевищує 3%. Різниця в накопиченні радіоактивного йоду та технеція, який не включається до складу тиреоїдних гормонів за 2 години, дає можливість встановити кількість йоду, що входить лише до органічної фракції, тобто досліджувати органічну фазу внутрішньотиреоїдного обміну йоду.
Дослідження транспортно-органічної фази йодного обміну (див.) здійснюють в основному за допомогою визначення концентрації тиреоїдних гормонів та тироксинзв'язуючого глобуліну в плазмі крові in vitro радіоімунологічним методом. Цей метод діагностики дозволяє з високим ступенем точності біологічно проаналізувати важливі компоненти, що залучаються до патологічного процесу. При цьому повністю виключається променеве навантаження на хворого.

Методи оцінки структури щитовидної залози включають комп'ютерну томографію (див. Томографія комп'ютерна), ехографію (див. Ультразвукова діагностика), радіонуклідне сканування і сцинтиграфію, пункційну біопсію, а також ряд спеціальних рентгенологічних методів рентгенотиреографію (див. Рентгенографія), електрорентгенотиреографію (див. Електрорентгенографія), тиреоїдолімфографію (рис. 3), пневмотиреографію, ангіотиреографію (див. Ангіографія). Впровадження комп'ютерної томографії, ехографії, радіонуклідного сканування та сцинтиграфії призвело до того, що спеціальні рентгенологічні методи втрачають своє значення.
Комп'ютерна томографія дозволяє отримати зображення щитовидної залози та навколишніх тканин. Нормальна щитовидна залоза на поперечних томограмах має вигляд двох однорідних за структурою овалів з відносно рівними добре відокремленими від навколишніх тканин контурами. При вузлових утвореннях у щитовидній залозі її структура виглядає гетерогенною. Контури утворень при вузловому зобі та раку щитовидної залози, як правило, менш чіткі, ніж при доброякісних пухлинах (аденомі, кісті та ін.). При пальпованої злоякісної пухлини комп'ютерна томографія дозволяє визначити форму, розміри, контури, структуру вузла, наявність та поширеність метастазів, а також ступінь залучення до патологічного процесу судин шиї та сусідніх тканин. Використання комп'ютерної томографії для діагностики вузлових новоутвореньта дифузних патологічних процесів щитовидної залози доцільно поєднувати з радіоімунологічними тестами, ультразвуковим та радіонуклідним скануванням.

Радіонуклідна тиреографія (сканування та сцинтиграфія) займає важливе місце в комплексному обстеженніхворих із патологією щитовидної залози. За допомогою цього методу оцінюють топографію щитовидної залози, її розміри, характер накопичення радіонукліду у різних ділянках залози. Хворому дають внутрішньо 0,025-0,05 мкюрі (1 - 2 МБк) 131I або 1,5-2,5 мкюрі (60 - 100 МБк) 99m Tc-пертехнетату і проводять дослідження через 2 і 24 години. У нормі на сканограмі чітко виділяються обриси щитовидної залози, її частки та перешийок. Максимальна радіоактивність посідає центр часток, до периферії часток інтенсивність випромінювання поступово падає і потім різко обривається. Розміри часток, їх форма дуже варіабельні. Пірамідальна частка найчастіше не виявляється. За допомогою цього способу легко виявляються різні аномалії положення органу. При дифузних формах тиреотоксичного зоба на сканограмі відзначається збільшене зображення щитовидної залози з інтенсивним рівномірним розподілом радіонукліду. В інших випадках (при хронічному тиреоїдиті, змішаному зобі) спостерігається нерівномірний розподіл радіонукліду. Сканування та сцинтиграфія дають можливість оцінити функціональний стан вузлів, які виявляються в тканині щитовидної залози, що має важливе значення для вибору тактики лікування. Так, морфологічним субстратом «гарячого» вузла найчастіше буває токсична аденома чи неавтономна гіперплазія тканини щитовидної залози (рис. 4, а). «Холодний» вузол є ділянкою нефункціонуючої тканини, кісту, аденому, метастаз пухлини (рис. 4, б). (Рис. 4, а). «Холодний» вузол є ділянкою нефункціонуючої тканини, кісту, аденому, метастаз пухлини (рис. 4, б).
За допомогою одновимірної та двовимірної ехографії (ультразвукового сканування) можна отримати інформацію про розміри щитовидної залози та її окремих ділянок. У нормі на ехограмі добре виявляються межі шкіри, підшкірної клітковини, фасцій, часток щитовидної залози, судин, м'язів, трахеї та хребта. При дифузному зобі зображення щитовидної залози не змінено, та її розміри збільшено. При хронічному тиреоїдиті та змішаному зобі відзначається зміна розмірів щитовидної залози та осередково-дифузна акустична неоднорідність зображення залози при нормальній картині навколишніх тканин, якщо трахея не зміщена. Вузловий зоб характеризується специфічною картиною, яка залежить від структури вузла. Зазвичай щільні вузли, аденоми, ділянки кальцинозу та кісти чітко визначаються на тлі незміненої тканини щитовидної залози. При раку щитовидної залози ехографічна картина залежить від характеру та поширеності патологічного процесу. При локальному розташуванні пухлини чи її метастазів вони можуть відрізнятися від щільних вузлів чи аденоми. При залученні до процесу суміжних тканин у них виявляються осередки ущільнення та тяжі. Ехографія у поєднанні з радіонуклідним скануванням дозволяє в більшості випадків встановити розміри та структуру щитовидної залози та її пухлин, що має важливе значення при виборі методу та обсягу оперативного втручання.
Пункція щитовидної залози тонкою голкою (пункційна біопсія), що здійснюється з діагностичною метою, може проводитись в амбулаторних умовах. Достовірність морфологічного діагнозу залежить від точності попадання голки в досліджувану ділянку, тому застосовують так звану придельную біопсію, яку проводять під контролем ехографії, або за даними радіонуклідного сканування.
У діагностиці захворювань щитовидної залози велике значення мають функціональні проби (тести), які проводяться шляхом введення трийодтироніну, тиреотропного гормону та тироліберину (рифатироїну). Тест придушення функції щитовидної залози (інгібіційний тест) використовують при діагностиці стертих форм тиреотоксикозу (див.), ендемічного зоба (див. Зоб ендемічний) та при диференціальній діагностиці офтальмопатій. Для цього спочатку проводять дослідження накопичення умов. Достовірність морфологічного діагнозу залежить від точності попадання голки в досліджувану ділянку, тому застосовують так звану придельную біопсію, яку проводять під контролем ехографії, або за даними радіонуклідного сканування.
Тест стимуляції функції щитовидної залози використовують для діагностики первинного та вторинного гіпотиреозу та функції виявлених у залозі вузлів. Визначають вміст тироксину в сироватці крові, після чого внутрішньом'язово вводять тиреотропний гормон, а потім радіонуклід (радіоактивний йод) з подальшим визначенням тироксину та дослідженням накопичення щитовидної залози радіоактивного йоду. У здорових осіб накопичення радіоактивного йоду щитовидною залозою або вміст тироксину в крові перевищують вихідні дані більш ніж на 20%. При первинному гіпотиреозі реакція на тиреотропний гормон відсутня. За наявності протипоказань для проведення радіонуклідного дослідження застосовують метод визначення тироксину у сироватці крові до введення тиреотропного гормону та через 24 години після його введення.
Тест стимуляції гіпофіза використовують для диференціальної діагностики різних видів гіпотиреозу. При цьому визначають вихідний рівень тиреотропного гормону у сироватці крові, потім вводять тіроліберин (внутрішньовенно або per os), після чого повторно визначають рівень тиреотропного гормону у сироватці крові. У здорових людей та при первинному гіпотиреозі рівень тиреотропного гормону в порівнянні з вихідним значно підвищується. При вторинному (гіпофізарному) гіпотиреозі та дифузному токсичному зобі реакція на тироліберин відсутня. Якщо у хворого збережена реакція на екзогенний тиреотропний гормон та тироліберин, слід думати про третинний (гіпоталамічний) гіпотиреоз.
Згідно з класифікацією, прийнятою в 1961 році на Міжнародному конгресісоціалістичних країн з проблеми ендемічного зоба, що виділяють вроджені аномаліїщитовидної залози, ендемічний зоб (і ендемічний кретинізм), спорадичний зоб, дифузний токсичний зоб, гіпотиреоз, запальні захворювання щитовидної залози (неспецифічні та специфічні), ушкодження та пухлини.
Вкрай рідко зустрічається аплазія щитовидної залози, причиною якої є порушення диференціювання ембріонального зачатку в тканину щитовидної залози. Аплазія щитовидної залози виявляється у ранньому дитячому віці. Гіпоплазія щитовидної залози обумовлена ??нестачею йоду в організмі матері. Клінічно у своїй спостерігається кретинізм (див.). Основним видом лікування є замісна терапія, яку призначають відразу після встановлення діагнозу і навіть у разі передбачуваного гіпотиреозу (див.). Своєчасне лікування може забезпечити нормальний фізичний розвиток дитини.
Збереження щитомовної протоки часто призводить до утворення серединних кіст та свищів шиї, зоба кореня язика. Свищі та кісти щитомовної протоки зазвичай розпізнаються у перші десять років життя дитини. Лікування - повне висічення кіст. Прогноз сприятливий.
Зміщення медіального зачатка щитовидної залози в середостіння зумовлює розвиток внутрішньогрудинного зоба (див. Середовище). Аномалія медіального зачатка щитовидної залози обумовлює дистопію тиреоїдної тканини в стінку трахеї, глотки, міокард, перикард, жирову клітковину середостіння, скелетні м'язишиї. Дистоповані вогнища тиреоїдної тканини можуть бути джерелом пухлин щитовидної залози. Виявлення тиреоїдної тканини у лімфатичних вузлах шиї розглядається як метастаз диференційованого раку щитовидної залози (див. нижче розділ Пухлини). За наявності зоба або пухлини у дистопованій тиреоїдній тканині показано оперативне лікування.
Закриті ушкодження щитовидної залози зустрічаються рідко (наприклад, здавлення шиї зашморгом при суїцидальній спробі) і виявляються утворенням гематоми. Показаний спокій та місцеве застосуванняхолоду. При наростанні гематоми, утрудненні дихання вдаються до зупинки кровотечі, а при необхідності – до трахеостомії (див.).
Відкриті ушкодження щитовидної залози зазвичай поєднуються з пораненням інших органів шиї (див.) та супроводжуються рясною кровотечею(Див.). У подібних випадках необхідні невідкладна хірургічна обробка рани з економною резекцією пошкодженої частини залози, зупинка кровотечі, ушивання ран з залишенням дренажу. Прогноз залежить від обсягу пошкодження.
Захворювання можуть протікати з ознаками підвищення щитовидної залози (тиреотоксикоз) або зниження її функції (гіпотиреоз). При деяких захворюваннях щитовидної залози порушення її функцій клінічно не виявляються (див. Еутиреоз).
Найбільш поширеним захворюванням щитовидної залози є ендемічний зоб (див. Зоб ендемічний), що зустрічається в географічних районахз недостатнім вмістом йоду в навколишньому середовищі. Захворювання супроводжується дифузним, вузловим чи змішаним збільшенням залози, здебільшого без порушення її функції. Причиною розвитку захворювання є недостатність йоду в організмі. При профілактичному застосуваннійодованою кухонної соліі препаратів йоду захворюваність населення різко скорочується
Зоб без виражених порушень функції щитовидної залози у осіб, які проживають у неендемічних районах, отримав назву спорадичного зобу (див. Зоб спорадичний).
Дифузне збільшення щитовидної залози з її гіперфункцією, що викликає порушення обміну речовин та розвиток патологічних змін у різних органах та системах, отримало назву «токсичний зоб». Розрізняють дифузний, вузловий та змішаний токсичний зоб (див. Зоб дифузний токсичний).
Зниження функції щитовидної залози-гіпотиреоз (див.) виникає в результаті ураження безпосередньо щитовидної залози (первинний гіпотиреоз), ураження гіпофіза (вторинний, або гіпофізарний, гіпотиреоз) або гіпоталамуса (третинний, або гіпоталамічний, гіпотиреоз).
До запальних захворювань щитовидної залози належать неспецифічні та специфічні (туберкульозний, сифілітичний, актиномікозний) тиреоїдити (див.). Розрізняють гострий, підгострий та хронічний тиреоїдит. Специфічні тиреоїдити зустрічаються дуже рідко і зазвичай є локальними проявами системних захворювань.
Пухлини частіше виникають на тлі посиленої тиреотропної функції гіпофіза, що зумовлює проліферацію епітелію щитовидної залози. Стимуляція тиреотропної функції гіпофіза може бути викликана аліментарною йодною недостатністю, антитиреоїдними засобами, впливом іонізуючого випромінювання (зовнішнє та внутрішнє опромінення), дисгормональними порушеннями. Розрізняють доброякісні та злоякісні пухлини щитовидної залози.
Доброякісні пухлини. Серед доброякісних пухлинчастіше зустрічаються аденоми (див. Аденома), зазвичай поодинокі, рідше множинні ( багатовузловий зоб), складові, за даними Слоуна та Франца (L. Sloan, W. Franz), 16% від усіх вузлових утворень щитовидної залози. Рідко спостерігаються фіброма, тератома, параганглиома, гемангіома, ліпома, міома.
За гістологічною будовою розрізняють трабекулярну (ембріональну), тубулярну (фетальну), мікрофолікулярну та макрофолікулярну (колоїдну) аденоми. Множинні аденоми щитовидної залози можуть мати різну будову та різну функціональну активність.
Аденоми, що не перевищують 1 см у діаметрі, клінічно не виявляються. Пухлина більшого розміру визначається у вигляді округлого безболісного вузла з гладкою поверхнею, що рухається при ковтанні. У міру зростання та при локалізації за грудиною аденома може здавлювати стравохід, трахею, викликаючи задишку (див.), рідше – дисфагію (див.).
У хворих з аденомами щитовидної залози функцію залози частіше не порушено (див. Еутиреоз). При токсичної аденомірозвиваються явища тиреотоксикозу (див.).
Трабекулярна та тубулярна аденоми не захоплюють радіоактивний йод. Аденоми, що мають фолікулярну будову, здатні захоплювати йод різною мірою та синтезувати тиреоїдні гормони.
Здатність аденоми захоплювати йод визначають за допомогою сканування щитовидної залози. Аденоми, що не захоплюють або слабо захоплюють радіоактивний йод, виявляються як «холодні» вузли, а аденоми, що активно захоплюють радіоактивний йод,- як «теплі» або «гарячі» вузли.
В аденомах можуть зустрічатися клітини. Пухлина, що повністю складається з цих клітин, іноді розглядають як великоклітинну онкоцитарну аденому. Такі аденоми найчастіше мономорфні, мають солідну та фолікулярно-солідну будову. Не виключена можливість їхнього інвазивного зростання.
Пухлини, подібні до фолікулярних аденом, але містять різна кількістьпапілярних (сосочкових) структур, деякі дослідники відносять до злоякісних. Питання можливості доброякісного варіанту медулярної пухлини (аденоми з парафоликулярных клітин) остаточно не вирішено.
Діагноз встановлюють на підставі даних кохмплексного обстеження хворих, що включає клініко-лабораторні, радіонуклідні, рентгенологічні методи та ін. Провідну роль у постановці діагнозу грає пункція пухлини щитовидної залози тонкою голкою (пункційна біопсія) з наступним цитологічно. У деяких випадках виникає необхідність проведення термінового гістологічного дослідження пухлини під час операції (інтраопераційна цитодіагностика).
Лікування доброякісних пухлин щитовидної залози оперативне. Операція полягає в резекції або повному видаленні ураженої частки залози (гемітиреоїдектомія). Широко поширена раніше операція енуклеації пухлини нині не застосовується.
Прогноз при радикальному лікуванні найчастіше сприятливий.
Злоякісні пухлини.За даними А. І. Пачеса та Р. М. Пропп (1984), рак становить понад 90% від усіх злоякісних пухлин щитовидної залози. Неепітеліальні пухлини, такі як саркома, злоякісна лімфома, гемангіоендотеліома, злоякісна тератома, в щитовидній залозі зустрічаються рідко. За будовою та клінічному перебігувони не відрізняються від аналогічних пухлин інших органів.
Рак щитовидної залози найчастіше зустрічається у жінок віком 40-60 років. Нерідко він розвивається на тлі тривалого, звичайно вузлового зоба (див. Передпухлинні захворювання), проте можливий розвиток раку (див.) і в незміненій залозі, рідко - на тлі дифузного токсичного зоба. Питання про зв'язок раку щитовидної залози ендемічним зобомостаточно не вирішено. Є дані про онкогенну роль рентгенівського опромінення області голови та шиї у дитячому та юнацькому віці.
Розрізняють диференційований та недиференційований рак щитовидної залози. Проміжне положення між ними займає медулярний рак. Крім того, у щитовидній залозі зустрічаються злоякісні пухлини з метаплазованого епітелію (плоскоклітинний рак).
До групи диференційованих пухлин щитовидної залози входять папілярний та фолікулярний рак. Папілярний рак ( папілярна аденокарцинома) - Найбільш часто зустрічається (близько 65%) форма раку щитовидної залози. Макроскопічно пухлина представлена частково інкапсульованим округлим або неправильною формою вузлом. Розміри пухлини значно варіюють. Вона може бути дуже маленькою (виявлятися тільки при мікроскопічному дослідженні) або займати всю залозу і поширюватися на навколишні тканини та органи. При мікроскопічному дослідженні виявляються характерні папілярні (сосочкові) структури, що становлять основну масу пухлини, та кістозні порожнини, заповнені колоїдом або кров'ю. Поряд з папілярними пухлини можуть зустрічатися фолікулярні структури, а в деяких випадках - солідні клітинні поля. Характерною ознакою папілярного раку щитовидної залози є осередкове відкладення солей кальцію у вигляді псаммозних тілець (див.).
Папілярний рак характеризується здатністю до інфільтруючого зростання з проростанням у капсулу щитовидної залози, лімфатичні і, рідше, кровоносні судини. Однією з типових ознак папілярного раку є метастазування в регіонарні лімфатичні вузли.
Розвиток пухлини повільний. Папілярний рак зазвичай функціонально мало активний і супроводжується ендокринними розладами.
Фолікулярний рак (фолікулярна аденокарцинома) спостерігається рідше за папілярний. Макроскопічно є досить добре відмежований вузол різних розмірів. Вузол малої величини нерідко виявляється випадково при гістологічному дослідженні тканини щитовидної залози, видаленої з іншого приводу, або проявляється клінічно метастазами в лімфатичних вузлах шиї, легенях і кістках. Мікроскопічно фолікулярний рак представлений фолікулярними та трабекулярними структурами, а також солідними розростаннями пухлинних клітин. Клітини фолікулярного раку можуть нагадувати тироцити нормальної щитовидної залози. Пухлина, що складається з високодиференційованих фолікулів, що містять колоїд, менш злоякісна, ніж пухлина, в якій переважають дрібні фолікули, що не містять колоїд, трабекулярні і особливо солідні структури.
Фолікулярний рак важко морфологічно диференціювати з фолікулярною аденомою. Інвазія пухлинних клітин у судини та капсулу щитовидної залози або наявність емболів з пухлинних клітин у кровоносних та лімфатичних судинах дозволяють поставити діагноз раку щитовидної залози.

Фолікулярний рак розвивається повільно, пухлина часто функціонально активна. Характерною ознакою є гематогенне метастазування, при якому насамперед уражаються легені (рис. 5) та кістки.
Різновидом папілярного, а іноді і фолікулярного раку щитовидної залози є так званий прихований рак, або мікрокарцинома, що склерозується.
Пухлина має дуже маленькі розміри, як правило, сосочкову будову з вираженими явищами склерозу. Метастази в регіонарні лімфатичні вузли шиї, які раніше помилково розцінювали як пухлини бічних аберантних щитовидних залоз, часто бувають єдиним клінічним проявом цього виду раку щитовидної залози.
Недиференційований рак щитовидної залози – одна з найбільш злоякісних пухлин людини; він становить 5-20% від усіх видів раку щитовидної залози. Макроскопічно пухлина найчастіше складається з декількох вузлів, що часто зливаються, не мають чітких меж. Пухлина щільна, на розрізі білуватого кольору, захоплює зазвичай всю щитовидну залозу, функціонально неактивна. Мікроскопічна картина недиференційованого раку щитовидної залози є неоднорідною. Пухлина може складатися з дрібних та гігантських поліморфних або веретеноподібних клітин. Нерідко в одній пухлині виявляють усі перелічені видиклітин, які ростуть суцільними клітинними полями та не утворюють фолікулярних чи папілярних структур.
Характерно бурхливий розвиток первинної пухлини та генералізоване метастазування. Пухлина проростає м'які тканини шиї, трахею, стравохід, зворотний гортанний нерв і судинно-нервовий пучок шиї. Тяжкими ускладненнями є стравохідно-трахеальні свищі (див. Бронхи, таблиця), асфіксія (див.) і кровотеча (див.) з судин пухлини, що розпадається.
Медулярний рак (рак з парафолікулярних клітин) становить 2-4% від усіх видів раку щитовидної залози. У ряді випадків пухлина генетично детермінована, поєднується з феохромоцитомою (див. Хромаффінома) та іншими хворобами ендокринної системи. Розвитку медулярного раку частіше передує осередкова гіперплазія парафолікулярних клітин. Макроскопічно медулярний рак представлений щільним пухлинним вузлом без чітких меж, який може бути як мікроскопічних розмірів (мікрокарцинома), так і займати всю щитовидну залозу та поширюватись за її межі. Пухлина рідко інкапсульована, часто впроваджується в тканину щитовидної залози, інфільтрує її капсулу та стінки. кровоносних судин. Гістологічна картина медулярного раку щитовидної залози є неоднорідною. Клітини переважно дрібні, округлі або витягнуті, можуть траплятися веретеноподібні клітини. Найчастіше у тканини медулярного раку визначається амілоїд. При електронній мікроскопії в пухлинних клітинахмедулярного раку, як і в нормальних парафолікулярних клітинах, виявляються характерні секреторні гранули та фібрилярні структури.
Пухлина гормонально-активна, продукує кал'цитонін (див.). Одним з характерних ознакмедулярного раку щитовидної залози є діарея, обумовлена впливом гуморальних факторів, секретованих пухлиною (кальцитоніну, серотоніну та ін). Медулярний рак характеризується відносно тривалим перебігом, частим метастазуванням в регіонарні лімфатичні вузли та рецидивуванням.
Плоскоклітинний (епідермоїдний) рак щитовидної залози становить 1-3% від усіх злоякісних пухлин щитовидної залози. Найчастіше спостерігається вторинне ураження щитовидної залози внаслідок поширення плоскоклітинного раку із сусідніх органів (гортані, стравоходу та ін.), а також при метастазах з інших органів. Ділянки плоскоклітинної метаплазії можуть зустрічатися в папілярному та фолікулярному раку. Пухлина може займати всю щитовидну залозу і поширюватися на навколишні тканини. Мікроскопічно пухлина має типову будову плоскоклітинного раку. Клин, течія вкрай важка, метастазування раннє і велике.
Поширеність раку щитовидної залози прийнято оцінювати за стадіями.
I стадія: невелика інкапсульована пухлина в одній із часток залози. II стадія: а) пухлина займає 1/2 залози, проростає її капсулу, рухлива; б) пухлина тих самих або менших розмірів з рухомими регіонарними метастазами на шиї з одного боку. III стадія: а) пухлина займає більше 1/2 або всю залозу, спаяна із сусідніми органами, обмежено рухлива; б) пухлина тих самих чи менших розмірів, але з двосторонніми метастазами в шийні лімфатичні вузли. IV стадія: а) пухлина проростає навколишні тканини та органи, нерухома; б) пухлина будь-яких розмірів, але з віддаленими метастазами.
Діагноз раку щитовидної залози у початкових стадіях важкий, оскільки інкапсульована ракова пухлинане має ознак, що дозволяють відрізнити її від аденоми. Використовують комплекс методів, серед яких провідна роль належить пункційної біопсії (див.), застосовують рентгенологічні (пневмотиреографія, артеріографія, тиреолімфографія, комп'ютерна томографія), радіонуклідні методи (див. Сканування, Сцинтиграфія), ехографія (див. Ультразвика). .). Лабораторні дані мають значення при медулярному раку, оскільки дозволяють визначити підвищену секрецію кальцитоніну. У сумнівних випадках показано оперативне втручання, обсяг якого залежить від результатів термічного гістологічного дослідження.
Основний метод лікування раку щитовидної залози – оперативне втручання. Операції при раку щитовидної залози роблять під ендотрахеальним наркозом (див. Інгаляційний наркоз). Уражену тканину видаляють екстра-капсулярно з перев'язкою судин протягом, виділенням зворотних гортанних нервів та паращитовидних залоз. У I стадії виробляють гемітиреоїдектомію з видаленням перешийка; у II стадії – субтотальну резекцію залози; у III та IV стадіях – тиреоїдектомію (див.). За наявності рухомих метастазів у регіонарні лімф, вузли поряд з тиреоїдектомією виробляють фасціально-футлярне висічення клітковини шиї з однієї або двох сторін. При обмежено зміщуваних метастазах у лімфатичних вузлах шиї з одного боку показано операцію Крайла (див. Крайла операція).
Як доповнення до оперативного методу при комбінованому лікуванні недиференційованого раку у передопераційному чи післяопераційному періоді використовують променеву терапію(Див.). При диференційованому раку променеву терапію призначають за неможливості здійснити радикальну операцію. Променева терапія пухлин щитовидної залози може застосовуватися і як самостійний виглядлікування або у комбінації з гормонотерапією при лікуванні неоперабельних первинних пухлин, метастазів у регіонарні лімфатичні вузли та віддалених метастазів.
У тих випадках, коли пухлина щитовидної залози та її метастази не накопичують або слабо накопичують I, променеву терапію здійснюють шляхом дистанційного опромінення. Лікування проводять на гамма-терапевтичних апаратах з джерелами 60 Co, 137 Cs або високоенергетичних прискорювачах з використанням гальмівного або електронного випромінювання (див. Гамма-терапія), а також шляхом прийому внутрішньо радіофармацевтичного препарату, міченого 131I, вибірково накопичується в тканині пухлинах з фолікулярного епітелію, що зберігають йодпоглинаючу функцію.
Для передопераційного опромінення рекомендуються сумарні дози 3000-4000 рад (30-40 Гр), для післяопераційного - 4000-5000 рад (40-50 Гр). У зону опромінення включають: область щитовидної залози, зони судинно-нервових пучків шиї та передньоверхнє середостіння. Для лікування неоперабельних пухлин та метастазів рекомендується сумарна доза не менше 6000 рад (60 Гр).
131I використовується головним чином для лікування віддалених метастазів, неоперабельних первинних пухлин і регіонарних метастазів, що мають йодпоглинаючу функцію. Лікування радіоактивним йодом проводиться до повного припинення йоднакопичувальних функцій у метастазах.
Гормонотерапія показана після радикального лікування як замісна терапія, а також для придушення продукції тиреотропного гормону гіпофіза з метою профілактики рецидиву та метастазів. Гормонотерапію проводять під контролем вмісту у крові тиреоїдних гормонів та тиреотропного гормону гіпофіза.
До сучасних протипухлинних препаратів рак щитовидної залози є резистентним. При поширеному процесі нетривалий ефект отримано при лікуванні дийодбензотефом, адріаміцином.
Прогноз залежить від стадії, гістологічної будови пухлини, статі та віку хворих. За даними Всесоюзного онкологічного наукового центру АМН СРСР, серед радикально лікованих хворих на рак щитовидної залози 5-річне виживання становило 90%, 10-річне - 86,4%.
Оперативне втручання на щитовидну залозу передбачає повне її видалення – тиреоїдектомію (див.) або часткове – резекцію щитовидної залози. У свою чергу, резекція щитовидної залози може полягати у видаленні частки залози (гемітиреоїдектомія) або субтотальної резекції щитовидної залози з залишенням 4-8 г її тканини. Показанням до оперативного втручання на щитовидній залозі є пухлини щитовидної залози, що тривалий час протікає хронічний тиреоїдит (див.), дифузний токсичний зоб (див. Зоб дифузний токсичний), а в деяких випадках - вузловий зоб (Зоб ендемічний). Абсолютних протипоказань до оперативного втручання на щитовидну залозу немає.
Операції на щитовидній залозі проводять під місцевою анестезієючи під ендотрахеальним наркозом. Вибір методу знеболювання індивідуальний і залежить від обсягу, технічної складності передбачуваної операції, віку та стану хворого.
Хворі з вузловим та дифузним зобом, що знаходяться в еутиреоїдному стані, спеціальної підготовки перед операцією не потребують. При тиреотоксичному зобі необхідна передопераційна підготовка з метою компенсації порушень, зумовлених тиреотоксикозом, та досягнення еутиреоїдного стану, що є профілактикою тиреотоксичного кризу у післяопераційному періоді (див. Зоб дифузний токсичний).
Комплекс засобів, що застосовуються для передопераційної підготовки, включає антитиреоїдні засоби (див.), Кортикостероїди (див.), А також засоби, що нормалізують серцеву діяльність, гіпотензивні, седативні засоби (див. Гіпотензивні засоби, седативні засоби). Для премедикації призначають також антигістамінні засоби (піпольфен) та промедол.
Можливі ускладнення, що виникають безпосередньо після операції, можуть бути: парез зворотного гортанного нерва, кровотеча, асфіксія; Незабаром після операції може виникнути тиреотоксичний криз (див. Кризи), гіпопаратиреоз, гіпотиреоз. У разі повного видалення щитовидної залози для профілактики гіпотиреозу необхідна замісна терапія, яка призначається незабаром після операції.
Ксенотрансплантація щитовидної залози при гіпотиреозі не застосовується через малу її ефективність; аутотрансплантація можлива за збереження віддаленої щитовидної залози у спеціальних умовах (див. Трансплантація).
Бібліогр.:Альошин Б. В. Про деякі спірні питання сучасної цитофізіології щитовидної залози, Усп. суч. біол., т. 93, ст. 1, с. 121, 1982; 0 н ж е, Проблема нейроендокринних клітин та гіпотези «дифузної ендокринної системи», там же, т. 98, ст. 1, с. 116, 1984: Альошин Би. Ст і Гускін В. І. Гіпоталамус і щитовидна залоза, М., 1983; Бомаш Н. Ю. Морфологічна діагностика захворювань щитовидної залози, М., 1981; Бухман А. ІІ. Рентгенодіагностика в ендокринології, М., 1974; Гольбер Л. М. і Канд-Ро р Ст І. Тиреотоксичне серце, М., 1972; Г о л ь б е р JI. М. та ін. Патогенез рухових розладів при тиреотоксикозі, М., 1980; Міський р-н В. М. і Козирицький В. Г. Ультраструктура залоз ендокринної системи, Київ, 1978; Зубовський Р. А. і Павлов Би. Р. Скенування внутрішніх органів, М., 1973; Іваницька В. І. та Шантир В. І. Променеві методи діагностики та лікування раку щитовидної залози, Київ, 1981; Кляч-ко В. Р. Актуальні питання консервативного лікуваннятоксичного зоба, М., 1965; Кондаленко В. Ф., Калінін А. П. і Одінокова В. А. Ультраструктура щитовидної залози людини в нормі і при патології, Арх. патол., Т. 32, № 4, с. 25, 1970; Лін-денбратен Л. Д. і Наумов Л. Б. Методи рентгенологічного дослідження органів і систем людини, Ташкент, 1976; О р а е ц В. Д. та М і р-ходжаев А. X. Вибір оптимального методу при математичній діагностиці захворювань щитовидної залози, Пробл. ендокрин., Т. 24, № 2, с. 23, 1978; Р. А. І. і Пропп Р. М. Рак щитовидної залози, М., 1984; Раскін А. М. Аутоімунні процеси в патології щитовидної залози, Л., 1968; Посібник з клінічної ендокринології, під ред. В. Г. Баранова, с. 348, М., 1979; Славно у В. Н. Радіоізотоп-ні та радіоімунологічні дослідження функції ендокринних залоз, Київ, 1978; Струков А. І. та Сєров В. В. Патологічна анатомія, с. 26, М., 1979; Тиреоїдні гормони, під ред. Я. X. Туракулова, с. 131, Ташкент, 1972; Фізіологія ендокринної системи, за ред. В. Г. Баранова, с. 135, Л., 1979; Ендокринна терапія злоякісних пухлин, під ред. Б. А. Столла, пров. з англ., с. 401, М., 1976; Bernal J. a. Refetoff S. The action of thyroid hormone, Clin. Endocr., v. 6, p. 227, 1977; Chung С. T. a. o. Внутрішнє irradiation for malignant thyroid tumors, Radiology, v. 136, p. 753, 1980; Ендокрінологія і метаболізм, ed. by Ph. Felig a. o., p. 281, N. Y.- Philadelphia, 1984; F u j i m o-t про Y. Thyroid tumors, Asian med. J., v. 25, p. 911, 1982; F u j i t a H. Добре structure of thethyroid gland, Int. Rev. Cytol., v. 40, p. 197, 1975; Hormones in blood, ed. by С. H. Gray a. H. T. James, v. 1-3, L. a. o., 1979; Labhart A. Klinik der inneren Sekretion, B. u. a., 1971; M e n g W. Schilddriisenerkrankun-gen, Jena, 1978; Rocmans P. A. a. o. Hormonal secretion hyperactive thyroid cells is not secundary to apical phagocytosis, Endocrinology, v. 103, p. 1834, 1978; Third International thyroid symposium, Thyroid cancer, Acta endocr., suppl. 252, 1983; Thethyroid, ed. by S. C. Werner a* S. H. Ingbar, Hagerstown a. o., 1978; Thyroid cancer, ed. by W. Duncan, B., 1980; Thyroid gland, ed. by M. de Visscher, N. Y., 1980.
H. Т. Старкова; Б. В. Альошин (біохім., фізіол.), Ю. І. Бородін (ан., гіст., ембр.), М. Е. Бронштейн, Ст А. Одинокова (пат. ан.), Е. С Кисельова (рад.), М. Ф. Логачов (пед.), А. X. Мирходжаєв (мет. ісл.), Р. М. Пропп (онк.), А. А. Філатов (рад., рент. ).
щитовидну залозу, в глиб органу відходять сполучнотканинні перегородки, які утворюють строму органу, містять судини та нерви. Розподіл паренхіми на часточки неповний і заліза тому є псевдодольчастою. Структурною одиницею щитовидної залози є фолікул - замкнута бульбашка, стінка якого вистелена одношаровим (фолікулярним) епітелієм.Вони вистилають стінку фолікулів, у порожнинах яких розташовується колоїд. Кожен фолікул оточений густою мережею капілярів (рис.), у просвіт яких секретуються гормони щитовидної залози – тироксин та трийодтиронін. У клітинах розрізняють апікальну, бічну та базальну поверхні. Базальна поверхня клітин знаходиться у тісному контакті з кровоносними капілярами, тут у плазмолемі є рецептори до тиротропіну; на бічних поверхнях тироцитів є оперізувальні замикальні контакти, на апікальній поверхні клітин розташовується багато мікроворсинок, в апікальній частині клітин лежить апарат Гольджі, різні типи везикул (секреторні, облямовані, ендоцитозні з незрілим і зрілим тиреоглобуліном) , тиреопероксидаза.
У незміненій щитовидній залозі фолікули рівномірно розподілені по всій паренхімі. Внаслідок заповнення просвіту фолікулів колоїдом тканина щитовидної залози є структурою, що містить велику кількість позаклітинної рідини (її обсяг більш ніж у 20 разів перевищує об'єм, зайнятий клітинами). Залежно від функціонального стану щитовидної залози, тироцити можуть бути плоскими, кубічними або циліндричними (рис.). При низькій функціональній активності залози тироцити, як правило, плоскі, при високій - циліндричні (висота клітин пропорційна ступеню активності здійснюваних у них процесів).
Колоїд, що заповнює просвіти фолікулів, є гомогенною в'язкою рідиною, що забарвлюється гематоксилін-еозином в рожевий колір. Основну масу колоїду становить тиреоглобулін, що секретується тироцитами у просвіті фолікула. Спочатку в гранулярній ендоплазматичній мережі синтезується поліпептидний ланцюжок глікопротеїду, до якого приєднуються бічні вуглеводні ланцюги. Процес завершується в апараті Гольджі створенням глікопротеїду, який у вигляді гранул транспортується до апікального полюса клітин та шляхом еккринії виділяється в порожнину фолікула. Розрізняють незрілий (нейодований або частково йодований) та зрілий (повністю йодований) колоїд.
Між тироцитами та кровоносними капілярами знаходиться базальна мембрана, а також прошарки пухкої волокнистої сполучної тканини. У цитоплазмі тироцитів добре розвинена гранулярна ендоплазматична мережа; мітохондрії, лізосоми, фаголізосоми.
В-клітини (клітини Ашкеназі-Гюртля) - більші за тироцити, мають еозинофільну цитоплазму і округле центрально розташоване ядро.
У цитоплазмі цих клітин виявлено біогенні аміни, зокрема серотонін. Вперше В-клітини з'являються у віці 14-16 років. У великій кількості вони зустрічаються у людей віком 50-60 років.
Парафолікулярні або С-клітини (у російській транскрипції К-клітини) відрізняються від тироцитів відсутністю здатності поглинати йод. Вони забезпечують синтез кальцитоніну – гормону, що бере участь в обміні кальцію в організмі. С-клітини більші за тироцити, у складі фолікулів розташовані, як правило, одиночно. Їхня морфологія характерна для клітин, що синтезують білок на експорт (присутні шорстка ендоплазматична мережа, комплекс Гольджі, секреторні гранули, мітохондрії). На гістологічних препаратах щитовидної залози цитоплазма С-клітин виглядає світлішою за цитоплазму тироцитів, звідси їх назва - світлі клітини.
Поряд з фолікулами в щитовидній залозі розрізняють інтерфолікулярні острівці, утворені тироцитами А, В, С. Острівці мають значення в регенерації паренхіми щитовидної залози у разі, якщо поразка має великий характер і супроводжується загибеллю цілих фолікулів. При часткових ушкодженнях фолікулів регенерація здійснюється за рахунок тироцитів, розташованих базально у стінці фолікула. Завдяки останнім відбувається фізіологічна регенерація фолікулярного епітелію.
Існує два погляди на механізм утворення нових фолікулів. Згідно з одним, проліферація базальних тироцитів веде до утворення інтерфолікулярних острівців, з яких виникають нові фолікули; або утворюються складки та фрагментація фолікулів. Таким чином, фолікулоутворення відбувається під дією внутрішньофолікулярних сил. Відповідно до другого погляду фолікулогенез здійснюється позафолікулярними силами - шляхом фрагментації вихідних фолікулів сполучнотканинними тяжами.
Оскільки основним елементом тяжів є перифолікулярні гемокапіляри, здатність викликати структурні перебудови пов'язують із наявністю скорочувальних мікрофіламентів у цитоплазмі ендотеліоцитів. Очевидно, крім транспортної та обмінної функцій гемокапіляри здатні виконувати і морфогенетичні функції у процесі онтогенезу. Морфогенетичну активність гемокапілярів індукують вазотропні гормони С-клітин (серотонін). С-клітини відносяться до дифузної нейроендокринної системи (ДНЕС), елементи якої локалізовані практично у всіх органах. Звідси випливає, що через клітини системи ДНЕС, які також продукують вазотропні гормони, та інших органах може здійснюватися морфогенетична функція капілярів.
У щитовидній залозі поряд із С-клітинами є й тканинні базофіли – клітини із набагато більш потужним арсеналом вазотропних гормонів. Численними дослідженнями доведено їхню здатність впливати на кровотік.
Якщо на тканинному рівні основним компартментом щитовидної залози є фолікули, оточені базальними мембранами, то однією з передбачуваних органних одиниць щитовидної залози можуть бути мікрорайони, до складу яких входять фолікули, клітини С, гемокапіляри, тканинні базофіли.
Оболонка з фібробластів, як правило, оточує групу з 4-6 фолікулів. Ця група (мікрочастка) є органним компартментом залози.
На момент народження щитовидна залоза функціонально активна і структурно цілком диференційована. У новонароджених фолікули дрібні (60-70 мкм у діаметрі), у дорослих – до 250 мкм. Вони розвинений інтерфолікулярний епітелій, характерний високий показник мітотичної активності. Є значний розкид у ступені розвитку фолікулів та інтерфолікулярних клітин.
У перші два тижні після народження фолікули інтенсивно розвиваються, а до 6 місяців вони добре розвинені по всій залозі, до року сягають 100 мкм у діаметрі. У період статевого дозрівання відмічено посилення зростання паренхіми та строми залози, підвищення її активності. Відзначено інтенсивне виведення колоїду та збільшення висоти тироцитів, підвищення в них активності ферментів. Фолікули набувають неправильної форми.
У процесі старіння маса щитовидної залози знижується, зменшується сумарний обсяг фолікулів та збільшується маса сполучної тканини. Фолікули різні за величиною, деякі переростяні колоїдом. Зменшується висота тироцитів та їх мітотична активність, знижено еозинофілію колоїду. У залозі збільшується кількість лімфоцитів, що сприймається як прояв аутоімунних процесів. Ці зміни розвиваються одночасно з перебудовою капілярної мережі. Інтерфолікулярний епітелій майже повністю зникає, мітози вкрай рідкісні. С-клітини суттєвої структурної перебудови не зазнають.
Санкт-Петербурзький державний медичний університет імені академіка І.П. Павлова
Реферат на тему:
«Цитофізіологія С-клітин щитовидної залози»
Студентка 2-го курсу
лікувального факультету
233 групи Локоткова А.М
Вступ
Найбільшою з ендокринних залоз людини є щитовидна залоза, що виділяє йодовмісні гормони та кальцитонін. Таким чином, вона здійснює гормональне регулювання процесів життєдіяльності та робить важливим розуміння її фізіологічного призначеннята цитології її клітин, у тому числі парафолікулярних.
Найпоширеніша пухлина органів ендокринної системи – рак щитовидної залози, що становить 0,5 % серед усіх новоутворень у чоловіків та 1 % у жінок. Усі дослідники одностайно стверджують, що частота раку щитовидної залози останні кілька десятиліть неухильно зростає (Камардин Л.Н., Романчишен А.Ф. 1980; Валдина Є.А., 1993). За даними Зарідзе Д.Г. (1992), у Росії захворюваність на рак щитовидної залози склала у чоловіків 0,96 і у жінок 3,09 на 100 000.
У США щорічно хворіють на рак щитовидної залози 50 осіб на 1 млн. жителів, причому перший пік захворюваності падає на вік 30-34 роки та другий пік – 60 років. У Франції частота захворюваності до 40 років - 10 на 100 000 населення (Валдіна Є.А).
На медулярний рак ЩЗ, який походить з парафолікулярних С-клітин, припадає 9% всіх пухлин щитовидної залози.
З цієї точки зору, актуальність моєї теми виправдана, тому що для того, щоб правильно діагностувати і прогнозувати перебіг патології необхідно розуміти і знати норму.
І в результаті метою мого реферування стало розкриття цитофізіології С-клітин щитовидної залози, а також їх гістогенезу і пов'язане з ними захворювання.
Закладка та первинне диференціювання парафолікулярних клітин щитовидної залози людини
Закладка кальцитонінсекретуючих клітин походить з ентодермальної частини рудименту останньої глоткової кишені. Імуногістохімічно показано, що до вбудовування закладки в щитовидну залозу клітини не секретують кальцитонін. Секреторна активність парафолікулярних клітин починається на 9-10 тижні ембріонального розвиткулюдини. Перші клітини, що секретують кальцитонін, з'являються у щитовидній залозі у вигляді дифузної мережі. Така мережа секреторних клітин формується за рахунок міграції попередників кальцитонінсекретуючих клітин із зони вбудовування в щитовидну залозу.
Парафолікулярні клітини щитовидної або кальцитоніноцити виникають у ембріонів людини на початку 2-го місяця (5 тижнів) пренатального розвитку. Вони закладаються незалежно від щитовидної залози і вбудовуються до 6-го тижня ембріонального розвитку. Кальцітоніноцити відносяться до бранхіогенної групи залоз внутрішньої секреції і виникають як деривати глоткової ентодерми, розташованої за IV ковтковою кишенею. Однак існують уявлення, згідно з якими С-клітини є похідними клітин нервового гребеня, які мігрують із зони змикання нервової трубки.
Ділянка глотки, що містить зачаток парафолікулярних клітин, вперше з'являється у людського ембріона на 4,5-му тижні розвитку. На цьому етапі зачаток виглядає як парне вентральне випинання глоткової ентодерми. Він неоднорідний формою і витягнутий вздовж осі тіла зародка. У ростральній частині зачаток має вигляд трубки, стінки якої вистелені недиференційованими клітинами глоткової ентодерми. У каудальній частині закладки внутрішня порожнина розширена та з'єднується з перитонельною порожниною. Закладка залози залишається сполученою з глотковою ентодермою до 5-го тижня розвитку. Після 5-го тижня розвитку зачаток, що містить парафолікулярні клітини, відокремлюється від глоткової ентодерми та зміщується у вентральному напрямку. У цей період зачаток набуває кулястої форми, а внутрішня порожнина зникає. До кінця 6-го тижня розвитку, зачаток, що містить парафолікулярні клітини, підходить до закладки щитовидної залози, і С-клітини розсіюються в ній, розташовуючись як поза фолікулів, так і в їх стінці.
Закладка С-клітин людини є органом, гомологічним ультимобранхіальній залозі нижчих хребетних. Під час закладки та морфологічного диференціювання заліза організована як рудимент глоткової кишені, яка є у нижчих хребетних залишком останньої зябрової щілини. Як і найбільш примітивних акул і амфібій, зачаток парафолікулярних клітин людини має трубчасту структуру. Наявність порожнини всередині зачатку вказує на зв'язок між фолікулярною організацією ультимобранхіальної залози нижчих хребетних та парафолікулярних клітин людини.(2)
Цитологія С-клітин щитовидної залози
С-клітини називаються так за першою літерою латинського найменування їхнього секреторного продукту – кальцитоніну. С-клітини захоплюють з крові попередники амінів, декарбоксилюють їх до відповідного аміну і накопичують разом з кальцитоніном у гранулах. У зв'язку з цим клітини відносяться до APUD-системи (АПУД-система, дифузна нейроендокринна система). Висловлюють припущення, що С-клітини синтезують та виділяють у малих кількостях також соматостатин, речовину P та пептид, пов'язаний з кальцитоніновим геном.
С-клітини розташовуються частіше поблизу фолікулів, чому їх називали раніше парафолікулярними клітинами. Вони не завжди легко відрізняються від тиреоїдного епітелію при звичайних забарвленнях зрізів, хоча мають і світлішу цитоплазму, за що їх назвали ще «світлі клітини», і в 1,5-2 рази більші за фолікулярні клітини. Вони поліганальної чи злегка витягнутої форми. Ядра в них більша і світліша, з 1-2 щільними ядерцями. У тих випадках, коли вони розташовуються інтрафолікулярно, знаходяться між тироцитами і базальною мембраною і від колоїду відокремлені цитоплазмою фолікуляних клітин. Вони виявляються у вигляді невеликих скупчень або поодинці, розташовуючись у різних частинах частки щитовидної залози, але частіше в центральних відділах. Виведення гормону в периваскулярне місце здійснюється шляхом екзоцитозу. Тиреокальцитонін є антагоністом парагормону навколощитовидної залози і мають гіпокальціємічну дію. С-клітини чутливі до концентрації кальцію в крові. Морфологічна активність проявляється у їх дегрануляції. При тривалій гіперкальціємії спостерігається їхня гіперплазія. Для виявлення С-клітин на світлооптичному рівні найкраще застосовувати аргірофільні реакції Гримеліуса та Сев'є-Мунже, які дозволяють завдяки наявності аргірофільних гранул у цитоплазмі легко виявляти їх приблизно у 90% випадків.
В результаті, цитологічного аналізу популяції парафолікулярних клітин щитовидної залози, що проводився ярославським медичним інститутам в 1985 році, в ході якого була вивчена структура парафолікулярних С-клітин щурів-самців в період від 10 хвилин до 8 годин після внутрішньочеревної ін'єкції кальцію С-клітин, що знаходяться на різних стадіях секреторного циклу.
Виділено такі типи С-клітин:
1. Клітини, цитоплазма яких повністю заповнена аргірофільними гранулами. По інтенсивності грануляції можна виділити: а) сильногранульовані форми, в яких гранули розташовані близько один до одного на відстані в середньому не більше одного їх поперечника, у ряді випадків гранули, що щільно прилягають, зливаються, і їх контури стають нечітко помітними; б) среднегранульированные форми, гранули у яких чітко помітні і розташовуються в цитоплазмі трохи рідше – з відривом у середньому 1-2 їх поперечника.
Матеріал узятий із сайту www.hystology.ru
Щитовидна залоза утворюється з ентодермального епітелію непарного серединного виросту вентральної стінки передньої кишки. Епітеліальні клітини формують складну систему тяжів. З мезенхіми розвивається сполучна тканинаяка покриває зачаток зовні і вростає в нього. З матеріалу непарного ембріонального органу утворюються дві частини, з'єднані перешийком. Останній зберігається на все життя лише у великого рогатої худобита свиней.
Щитовидна залоза розташована в області шиї по обидва боки трахеї, позаду щитовидного хряща.
Зовні щитовидна залоза покрита сполучнотканинною капсулою, від якої в глиб органу відходять перегородки, що розділяють паренхіму органа на часточки, а часточки на замкнуті бульбашки - фолікули (рис. 226).
Основною морфофункціональною структурою щитовидної залози є фолікул - замкнутий округлий або овальний пляшечку. Розміри фолікулів варіюють від 0,02 до 0,9 мм у діаметрі. У фолікулі розрізняють стінку та порожнину, заповнену колоїдом. Стінка фолікула складається з одношарового епітелію, розташованого на базальній мембрані.
Мал. 226. Щитоподібна залоза коня:
1 - фолікул; 2 - Стінка фолікула; 3 - колоїд; 4 - Вакуоля; 5 – капіляр; 6 - сполучна тканина.
Форма клітин обумовлена функціональною активністю щитовидної залози і може бути або плоскою, або кубічною, або стовпчастою (циліндричною). Якщо заліза характеризується помірною функцією, клітини фолікула мають кубічну форму. При підвищеній діяльності залози (гіперфункція) відзначають посилене надходження гормону в кров, клітини набувають стовпчастої форми (див. цв. табл. VII - б).Зниження функціональної активності залози (гіпофункція) пов'язане зі збільшенням діаметра фолікулів та накопиченням колоїду в їх порожнинах. Різко зменшується у своїй висота клітин. Вони стають сплощеними (В).
Функціональний стан залози впливає і консистенцію колоїда. При помірній функції колоїд гомогенний та заповнює всю порожнину фолікула. При гіперфункції колоїд більш рідкої консистенції має пінистий вигляд, багато вакуолей; знижується вміст колоїду у фолікулах. При гіпофункції колоїд згущується та ущільнюється.
Внутрішня вистилка фолікулів представлена клітинами двох типів: фолікулярними клітинами (тироцитами) та навколофолікулярними клітинами (К-клітинами). Останні зустрічаються рідше і можуть розташовуватися у стінці фолікула, а й з-поміж них. Функція тироцитів зводиться до синтезу йодовмісних гормонів тироксину і трийодтироніну. Вони регулюють окислювальні процеси, що впливають попри всі види обміну речовин, які у організмі. Гормонообразовательная функція фолікулярних клітин стимулюється тиротропними гормонами, тому вони належать до групи ендокринних клітин, функція яких залежить від передньої частки гіпофіза.
Навколофолікулярні клітини виробляють йоднемісткий гормон - кальцитонін (тирокальцитонін), який знижує вміст кальцію в крові і є антагоністом паратгормону, що синтезує навколощитовидною залозою. Гормональна функція навколофолікулярних клітин (К-клітин) залежить від передньої частки гіпофіза.
Фолікулярні клітини мають світле, центрально розташоване округле ядро. У цитоплазмі базального полюса знаходяться добре розвинені мембранні структури гранулярної ендоплазматичної мережі, мітохондрії з незначною кількістю христ.
Плазмолема утворює базальну складчастість. Над ядром чи біля нього лежить комплекс Гольджі, лізосоми. У цитоплазмі є дрібні краплі колоїду. Плазмолема апікального полюса утворює мікроворсинки, що збільшують поверхню контакту тироцитів із порожниною фолікула. Між собою клітини з'єднуються за допомогою плям злипання та термінальних платівок.
Навколофолікулярні (світлі) клітини - К-клітини розташовані у стінці фолікулів або у складі міжфолікулярних острівців, що лежать у міжфолікулярній сполучній тканині. Це світлі, великі, овальні клітини, апікальна поверхня яких не контактує з порожниною та колоїдом фолікула. У К-клітинах добре розвинені гранулярна ендоплазматична мережа, комплекс Гольджі, що свідчить про інтенсивний синтез білків; цитоплазма містить білкові секреторні гранули 0,1 - 0,4 мкм у діаметрі, незначна кількість мітохондрій. Особливістю цих клітин є нездатність поглинати йод.
Складовими клітинами міжфолікулярних острівців є епітеліальні клітини, які виконують функцію джерела розвитку нових фолікулів.
Зовні фолікули вкриті базальною мембраною. Фолікули відмежовані тонкими прошарками пухкої сполучної тканини, інтенсивно забезпеченої гемо-і лімфосудинної мережею. Міжфолікулярна сполучна тканина, з'єднуючись з міждольковою, утворює строму органа.
Секреторна діяльність фолікулярних клітин (тироцитів) дуже складна та зводиться до наступного.
1. З амінокислот і солей, що приносяться з кров'ю та проникають у тироцит, за активної участі рибосом, ендоплазматичної мережі, комплексу Гольджі утворюється неіодований тиреоглобулін, однією з амінокислот якого є тирозин. У вигляді дрібних секреторних бульбашок він накопичується в апікальній зоні тироцитів і за допомогою екзоцитозу надходить у порожнину фолікула.
2. У порожнині фолікула до тирозину тиреоглобуліну включаються послідовно атоми йоду, які утворюються при окисленні йодиду, поглиненого з крові фолікулярними клітинами. У ході цього процесу послідовно синтезуються монойодтирозип, дийодтирозин, тетрайодтірозін (тироксин), трийодтиронін і накопичуються в колоїді.
3. Тироцити своєю апікальною поверхнею шляхом ендоцитозу поглинають (фагоцитують) ділянки інтрафолікулярного колоїду, які всередині цитоплазми перетворюються на внутрішньоклітинні краплі колоїду. Лізосоми з'єднуються з ними, після їх розщеплення утворюються тиреоїдні гормони. Через базальну частину тироциту та базальну мембрану вони надходять у загальний кровотік, або в лімфатичні судини (рис. 227, 228).
Таким чином, до складу гормонів, що продукуються тироцитами, обов'язково входить йод, тому для нормальної функції щитовидної залози необхідний постійний його приплив з кров'ю до

Мал. 227. Фолікулярна клітина щитовидної залози (електронна мікрофотографія):
А – апікальна частина клітини, звернена до поверхні; I – мікроворсинки; 2 - Апікальні гранули; Б- органели, що беруть участь у секреції тиреоглобуліну; 3 - Розтягнуті цистерни гранулярної ендоплазматичної мережі; 4 - комплекс Гольджі; 5 - Транспортні бульбашки; 6 - просекреторні гранули; 7 – секреторні гранули; 8 - облямовані бульбашки; 9 - лізосоми; 10 - Мітохондрії.

Мал. 228. Околофолікулярна клітина (електронна мікрофотографія):
1 - ядро; 2 - Секреторні гранули.
щитовидної залози. Йод отримує організм із водою та продуктами харчування.
Рясне кровопостачання щитовидної залози забезпечується сонною артерією. За ступенем кровопостачання щитовидна залоза займає одне з перших місць серед інших органів.
Щитовидна залоза іннервується нервовими волокнами симпатичної та парасимпатичної нервової системи.
Основною структурно-функціональною одиницею щитовидної залози є фолікули. Вони є округлими порожнинами, стінка яких утворена одним рядом клітин кубічного епітелію. Фолікули заповнені колоїдом і містять гормони тироксин та трийодтиронін, які пов'язані з білком тиреоглобуліном. У міжфолікулярному просторі проходять капіляри, що забезпечують рясну васкуляризацію фолікулів. У щитовидній залозі об'ємна швидкість кровотоку вища, ніж в інших органах та тканинах. У міжфолікулярному просторі знаходяться парафолікулярні клітини (С-клітини), в яких виробляється гормон тиреокальцитонін.
Біосинтез тироксину та трийодтироніну здійснюється за рахунок йодування амінокислоти тирозину, тому в щитовидній залозі відбувається активне поглинання йоду. Вміст йоду у фолікулах у 30 разів перевищує його концентрацію в крові, а при гіперфункції щитовидної залози це співвідношення стає ще більшим. Поглинання йоду здійснюється з допомогою активного транспорту. Після з'єднання тирозину, що входить до складу тиреоглобуліну, з атомарним йодом утворюються монойодтирозин та дийодтирозин. За рахунок з'єднання 2 молекул дийодтирозину утворюється тироксин; конденсація моно- та дийодтирозину призводить до утворення трийодтироніну. Надалі за рахунок дії протеаз, що розщеплюють тиреоглобулін, відбувається вивільнення до крові активних гормонів.
Активність тироксину в кілька разів менша, ніж трийодтироніну. Крім того, ефекти трийодтироніну мають менший. латентний періодтому його дія розвивається значно швидше. З іншого боку, вміст тироксину в крові приблизно в 20 разів більший, ніж трийодтироніну. Тироксин при дейодуванні може перетворюватися на трийодтиронін. З цих фактів припускають, що основним гормоном щитовидної залози є трийодтиронин, а тироксин виконує функцію його попередника.
Дія гормонів щитовидної залози проявляється різким посиленням метаболічної активності організму. При цьому прискорюються всі види обміну речовин (білковий, ліпідний, вуглеводний), що призводить до збільшення енергоутворення та підвищення основного обміну. У дитячому віці це має важливе значення для процесів росту, фізичного розвитку, а також енергетичного забезпечення дозрівання тканини мозку, тому нестача гормонів щитовидної залози у дітей призводить до затримки розумового та фізичного розвитку (кретинізм). У дорослих при гіпофункції щитовидної залози спостерігається гальмування нервово-психічної активності (млявість, сонливість, апатія); при надлишку гормонів, навпаки, спостерігаються емоційна лабільність, збудження, безсоння.
Внаслідок активізації всіх видів обміну речовин під впливом гормонів щитовидної залози змінюється діяльність практично всіх органів. Посилюється теплопродукція, що зумовлює підвищення температури тіла. Прискорюється робота серця (тахікардія, підвищення артеріального тиску, збільшення хвилинного об'єму крові), стимулюється діяльність травного тракту(Підвищення апетиту, посилення перистальтики кишечника, збільшення секреторної активності). При гіперфункції щитовидної залози, зазвичай, знижується маса тіла. Нестача гормонів щитовидної залози призводить до змін зворотного характеру.
Кальцитонін, або тиреокальцитонін, знижує рівень кальцію у крові. Він діє на кісткову систему, нирки та кишечник, викликаючи при цьому ефекти, протилежні дії паратирину. У кістковій тканині тиреокальцитонін посилює активність остеобластів та процеси мінералізації. У нирках та кишечнику пригнічує реабсорбцію кальцію та стимулює зворотне всмоктування фосфатів. Реалізація цих ефектів призводить до гіпокальціємії.
Секреція гормонів щитовидної залози регулюється гіпоталамічним тиреоліберином. Вироблення тироксину та трийодтироніну різко посилюється в умовах тривалого емоційного збудження. Відзначено також, що секреція цих гормонів прискорюється при зниженні температури тіла.

Біологічно активна добавка на основі екстракту, отриманого з жука шпанкою мушки (або шпанки...)

Катерина МірімановаСистема мінус 60. РеволюціяСистема мінус 60 з Катериною Мірімановою «Система мінус 60. Революція»...

Тяжкість і здуття є причинами як звичайного переїдання, так і більш серйозних проблем із травним трактом.

Друга група крові резус-негативний з'явилася вже дуже багато років тому, коли людина перестала бути мисливцем, і...

ПСИХОЛОГІЧНІ АСПЕКТИ ФЕНОМЕНА АНОРЕКСІЇ (ЕКСПЕРИМЕНТАЛЬНЕ ДОСЛІДЖЕННЯ) Т. В. Тарасова, Є. В. Арсентьєва У...

Вигляд того, що шкіра в цій області тонка, вона більш схильна до появи різного роду плям. Почервоніння під...

vseslav сб, 10/17/2015 - 20:50 Станція "Василеострівська" - входить до найстаріших станцій...

Найчастіше гриб козляк (Suillus bovinus) називається в повсякденному мовленні грибників козеням. Цей трубчастий...

Кожну людину хоч раз у житті відвідували сновидіння. Багато хто не надає їм жодного значення, але деякі...

Дедал (кратер). Діаметр: 93 км Глибина: 3 км (фото НАСА) Місяць привертав увагу людей із давніх часів. У ІІ...

У третій понеділок лютого у США відзначається Президентський день. До 70-х років святкування були присвячені...

Свого улюбленого завуча в день останнього дзвінка я хочу сказати лише одне слово: «Дякую». Але як багато це...

Кавун вважається ласою і корисною ягодою, особливо корисне його насіння. Маленькі, темно-коричневі або...

«Розвиток мови дітей 6 років» Дитина не народжується з мовою, що склалася. Не можна однозначно відповісти на запитання про...

Катерина МірімановаСистема мінус 60. РеволюціяСистема мінус 60 з Катериною Міріманової«Система мінус 60....
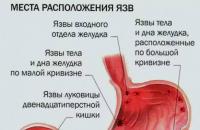
Тяжкість і здуття є причинами як звичайного переїдання, так і серйозніших проблем із травним...