




Благовещенский собор Московского Кремля
Один из древнейших храмов Московского Кремля стоит на краю Соборной площади на бровке Боровицкого холма. Много веков...
Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,
к окончанию телофазы
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой. М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять профаза, прометафаза, метафаза, анафаза и телофаза составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.

К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.

Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона около 30 минут.
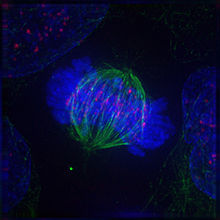
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.

Анафаза самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.

Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
| Прево, Жан-Луи |
Митоз, его фазы, биологическое значение
Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл - это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
Основные стадии митоза.
1.Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90% информации эукариотической клетки.
2.Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период).
Фазы клеточного цикла:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления;
2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка.
В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период);
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный период - препрофазу.
После этого наступает собственно митоз, который состоит из четырех фаз. Процесс деления включает в себя несколько последовательных фаз и представляет собой цикл. Его продолжительность различна и составляет у большинства клеток от 10 до 50 ч. При этом у клеток тела человека продолжительность самого митоза составляет 1-1,5 ч, G2-периода интерфазы - 2-3 ч, S-периода интерфазы - 6-10 ч.
Стадии митоза.
Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 1–3). Так как он непрерывен, смена фаз осуществляется плавно - одна незаметно переходит в другую.
В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть - прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n2хр).
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено - одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза
Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона - около 30 минут.
Метафаза
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи - к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Анафаза - самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза
Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки - цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру - раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
Среди всех интересных и достаточно сложных тем в биологии стоит выделить два процесса деления клеток в организме – мейоз и митоз . Сначала может показаться, что эти процессы одинаковые, поскольку в обоих случаях происходит деление клеток, но на самом деле между ними существует большая разница. В первую очередь, нужно разобраться с митозом. Что этот процесс из себя представляет, что такое интерфаза митоза и какую роль они играют в человеческом организме? Подробнее об этом и пойдет речь в данной статье.
Сложный биологический процесс, который сопровождается делением клеток и распределением хромосом между этими клетками – все это можно сказать о митозе. Благодаря ему, между дочерними клетками организма равномерно распределяются хромосомы, в которых содержится ДНК.

Существует 4 основные фазы процесса митоза. Все они связаны между собой, поскольку фазы плавно переходят из одной на другую. Распространенность митоза в природе обусловлена тем, что именно он участвует в процессе деления всех клеток, среди которых мышечные, нервные и так далее.

Перед попаданием в состояние митоза клетка, которая разделяется, переходит в период интерфазы, то есть растет. Длительность интерфазы может занимать более 90% всего времени активности клетки в обычном режиме .

Интерфаза делится на 3 основных периода:
Все они проходят в определенной последовательности. Рассмотрим каждую из этих фаз отдельно.

Интерфаза — основные составляющие (формула)
Этот период характеризуется подготовкой клетки к делению. Она увеличивается в объемах для дальнейшей фазы синтеза ДНК.
Это следующий этап в процессе интерфазы, при котором происходит деление клеток организма. Как правило, синтез большей части клеток происходит на небольшой промежуток времени. После деления клетки не увеличиваются в размерах, а начинается последняя фаза.

Финальный этап интерфазы, на протяжении которого клетки продолжают синтезировать белки, увеличиваясь при этом в размерах. В этот период в клетке по-прежнему есть нуклеолы. Также в последней части интерфазы происходит дублирование хромосом, а поверхность ядра в это время покрывается специальной оболочкой, имеющей защитную функцию.
На заметку! По завершению третьей фазы наступает митоз. Он тоже включает в себя несколько стадий, после которых происходит деление клетки (этот процесс в медицине называется цитокинезом).
Как уже отмечалось ранее, митоз делится на 4 стадии, но иногда их может быть и больше. Ниже представлены основные из них.
Таблица. Описание основных фаз митоза.
| Название фазы, фото | Описание |
|---|---|
| Во время профазы происходит спирализация хромосом, в результате чего они принимают скрученную форму (она более компактная). Останавливаются все синтетические процессы в клетке организма, поэтому рибосомы уже не вырабатываются. |
| Многие специалисты не выделяют прометафазу как отдельную фазу митоза. Нередко все процессы, которые в ней происходят, относят к профазе. В этот период цитоплазма окутывает хромосомы, которые свободно перемещаются по клетке до определенного момента. |
| Следующая фаза митоза, которая сопровождается распределением на экваториальной плоскости конденсированных хромосом. В этот период происходит обновление микротрубочек на постоянной основе. При метафазе хромосомы расположены так, что их кинетохоры находятся в ином направлении, то есть направлены к противоположным полюсам. |
| Данная фаза митоза сопровождается отделением хроматид каждой из хромосом друг от друга. Нарастание микротрубочек прекращается, они теперь начинают разбираться. Анафаза длится недолго, но за этот промежуток времени клетки успевают разойтись ближе к разным полюсам в примерно равном количестве. |
| Это последняя стадия, на протяжении которой начинается деконденсация хромосом. Эукариотические клетки завершают свое деление, а вокруг каждого набора хромосом человека образовывается специальная оболочка. При сокращении сократительного кольца происходит разделение цитоплазмы (в медицине этот процесс называется цитотомией). |
Важно! Длительность полного процесса митоза, как правило, составляет не больше 1,5-2 часов. Продолжительность может меняться в зависимости от вида разделяемой клетки. Также на длительность процесса влияют и внешние факторы, такие как световой режим, температура и так далее.

Теперь попробуем разобраться с особенностями митоза и его важностью в биологическом цикле. В первую очередь, он обеспечивает многие процессы жизнедеятельности организма, среди которых – эмбриональное развитие .

Также митоз отвечает за восстановление тканей и внутренних органов организма после различных видов повреждения, в результате чего происходит регенерация. В процессе функционирования клетки постепенно отмирают, но с помощью митоза структурная целостность тканей постоянно поддерживается.

Митоз обеспечивает сохранение определенного количества хромосом (оно соответствует числу хромосом в материнской клетке).
Мейоз - это деление в зоне созревания половых клеток , сопровождающееся уменьшением числа хромосом вдвое. Он состоит из двух последовательно идущих делений, имеющих те же фазы, что и митоз. Однако, как показано в таблице «Сравнение митоза и мейоза», продолжительность отдельных фаз и происходящие в них процессы значительно отличаются от процессов, происходящих при митозе.
Эти отличия в основном состоят в следующем.
В мейозе профаза I более продолжительна. В ней происходит конъюгация(соединение гомологичных хромосом) и обмен генетической информацией. В анафазе Iцентромеры, скрепляющие хроматиды, не делятся, а к полюсам отходит одна из гомологмейоза Митоз и его фазы митоза и ичных хромосом. Интерфаза перед вторым делением очень короткая, в ней ДНК не синтезируется. Клетки (галиты), образующиеся в результате двух мейотических делений, содержат гаплоидный (одинарный) набор хромосом. Диплоидность восстанавливается при слиянии двух клеток - материнской и отцовской. Оплодотворенную яйцеклетку называютзиготой.
Митоз, или непрямое деление, наиболее широко распространен в природе. Митоз лежит в основе деления всех неполовых клеток (эпителиальных, мышечных, нервных, костных и др.). Митоз состоит из четырех последовательных фаз (см. далее таблицу). Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними. Период жизни клетки между двумя митозами называют интерфазой. Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков , удваивается каждая хромосома, образуя две сестринские хроматиды, скрепленные общей центромерой, увеличивается число основных органоидов цитоплазмы.
В профазе спиралируются и вследствие этого утолщаются хромосомы, состоящие из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредоточиваются по всей клетке, центриоли отходят к полюсам и образуют веретено деления. В метафазе происходит дальнейшая спирализация хромосом. В эту фазу они наиболее хорошо видны. Их центромеры располагаются по экватору. К ним прикрепляются нити веретена деления.
В анафазе центромеры делятся, сестринские хроматиды отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки.
В телофазе цитоплазма делится, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. В животных клетках цитоплазма перешнуровывается, в растительных - в центре материнской клетки образуется перегородка. Так из одной исходной клетки (материнской) образуются две новые дочерние.
Мейоз и митоз
Таблица - Сравнение митоза и мейоза
|
1 деление |
2 деление |
||
|
Интерфаза |
Набор хромосом 2n Идет интенсивный синтез белков, АТФ и других органических веществ Удваиваются хромосомы, каждая оказывается состоящей из двух сестринских хроматид, скрепленных общей центромерой. |
Набор хромосом 2n Наблюдаются те же процессы, что и в митозе, но более продолжительна, особенно при образовании яйцеклеток. |
Набор хромосом гаплоидный (n). Синтез органических веществ отсутствует. |
|
Непродолжительна, происходит спирализация хромосом, исчезают ядерная оболочка, ядрышко, образуется веретено деления |
Более длительна. В начале фазы те же процессы, что и в митозе. Кроме того, происходит конъюгация хромосом, при которой гомологичные хромосомы сближаются по всей длине и скручиваются. При этом может происходить обмен генетической информацией (перекрест хромосом) -кроссинговер . Затем хромосомы расходятся. |
Короткая; те же процессы, что и в митозе, но при nхромосом. |
|
|
Метафаза |
Происходит дальнейшая спирализация хромосом, их центромеры располагаются по экватору. |
Происходят процессы, аналогичные тем, что и в митозе. | |
|
Центромеры, скрепляющие сестринские хроматиды, делятся, каждая из них становится новой хромосомой и отходит к противоположным полюсам. |
Центромеры не делятся. К противоположным полюсам отходит одна из гомологичных хромосом, состоящая из двух хроматид, скрепленных общей центромерой. |
Происходит то же, что и в митозе, но при nхромосом. |
|
|
Телофаза |
Делится цитоплазма, образуются две дочерние клетки, каждая с диплоидным набором хромосом. Исчезает веретено деления, формируются ядрышки. |
Длится недолго Гомологичные хромосомы попадают в разные клетки с гаплоидным набором хромосом. Цитоплазма делится не всегда. |
Делится цитоплазма. После двух мейотических делений образуется 4 клетки с гаплоидным набором хромосом. |
Клеточный цикл - это период существования клетки от момента её образования путем деления материнской клетки до собственного деления.
Длительность клеточного цикла эукариот
Длительность клеточного цикла у разных клеток варьируется. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12-36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих , земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10-24 ч.
Фазы клеточного цикла эукариот
Клеточный цикл эукариот состоит из двух периодов:
Период клеточного роста, называемый «интерфаза », во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Периода клеточного деления, называемый «фаза М» (от слова mitosis - митоз ).
Интерфаза состоит из нескольких периодов:
G 1 -фазы (от англ. gap - промежуток), или фазы начального роста , во время которой идет синтез мРНК , белков , других клеточных компонентов;
S-фазы (от англ. synthesis - синтез), во время которой идет репликация ДНК клеточного ядра , также происходит удвоение центриолей (если они, конечно, есть).
G 2 -фазы, во время которой идет подготовка к митозу .
У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G 1 фаза. Такие клетки находятся в фазе покоя G 0 .
Период клеточного деления (фаза М) включает две стадии:
-кариокинез (деление клеточного ядра);
-цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий.
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатахсветовой и электронной микроскопии фиксированных и окрашенных клеток.
Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков , какциклин-зависимые киназы и циклины . Клетки , находящиеся в G 0 фазе, могут вступать в клеточный цикл при действии на нихфакторов роста . Разные факторы роста, такие как тромбоцитарный , эпидермальный, фактор роста нервов, связываясь со своимирецепторами , запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов и циклин-зависимых киназ . Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами . Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса.Киназы не активны без циклинов . На разных стадиях клеточного цикла синтезируются разные циклины . Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза , когда запускается весь каскад реакций фосфорилирования , катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Контрольные точки клеточного цикла
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку, то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт. Существует как минимум четыре контрольных точки клеточного цикла: точка в G1, где проверяется интактность ДНК, перед вхождением в S-фазу, сверочная точка в S-фазе, в которой проверяется правильность репликации ДНК, сверочная точка в G2, в которой проверяются повреждения, пропущенные при прохождении предыдущих сверочных точек, либо полученные на последующих стадиях клеточного цикла. В G2 фазе детектируется полнота репликации ДНК, и клетки, в которых ДНК недореплицирована, не входят в митоз. В контрольной точке сборки веретена деления проверяется, все ли кинетохоры прикреплены к микротрубочкам.
Нарушения клеточного цикла и образование опухолей
Увеличение синтеза белка p53 ведет к индукции синтеза белка p21 - ингибитора клеточного цикла
Нарушение нормальной регуляции клеточного цикла является причиной появления большинства твердых опухолей. В клеточном цикле, как уже говорилось, прохождение контрольных пунктов его возможно только в случае нормального завершения предыдущих этапов и отсутствия поломок. Для опухолевых клеток характерны изменения компонентов сверочных точек клеточного цикла. При инактивации сверочных точек клеточного цикла наблюдается дисфункция некоторых опухолевых супрессоров и протоонкогенов, в частности p53 , pRb , Myc иRas . Белок p53 является одним из факторов транскрипции, который инициирует синтез белка p21 , являющегося ингибитором комплекса CDK-циклин, что приводит к остановке клеточного цикла в G1 и G2 периоде. Таким образом клетка, у которой повреждена ДНК, не вступает в S-фазу. При мутациях, приводящих к потере генов белка p53, или при их изменениях, блокады клеточного цикла не происходит, клетки вступают в митоз, что приводит к появлению мутантных клеток, большая часть из которых нежизнеспособна, другая - дает начало злокачественным клеткам.
Деление клеток
Все клетки появляются путём деления родительских клеток. Большинству клеток свойственен клеточный цикл, состоящий из двух основных стадий: интерфазы и митоза.
Интерфаза состоит из трех этапов. В течение 4–8 часов после рождения клетка увеличивает свою массу. Некоторые клетки (например, нервные клетки мозга) навсегда остаются в этой стадии, у других же в течение 6–9 часов удваивается хромосомная ДНК. Когда масса клетки увеличивается в два раза, начинается митоз .
В стадии анафазы хромосомы перемещаются к полюсам клетки. Когда хромосомы достигают полюсов, начинается телофаза . Клетка делится надвое в экваториальной плоскости, нити веретена разрушаются, вокруг хромосом формируются ядерные мембраны. Каждая дочерняя клетка получает собственный набор хромосом и возвращается в стадию интерфазы. Весь процесс занимает около часа.
Процесс митоза может варьировать в зависимости от типа клетки. В растительной клетке отсутствуют центриоли, хотя веретено деления образуется. В грибных клетках митоз происходит внутри ядра, ядерная мембрана не распадается.
Наличие хромосом не является необходимым условием деления клетки. С другой стороны, один или несколько митозов могут останавливаться на стадии телофазы, в результате чего возникают многоядерные клетки (например, у некоторых водорослей).
Размножение при помощи митоза называют бесполым или вегетативным, а также клонированием . При митозе генетический материал родительских и дочерних клеток идентичен.
Мейоз , в отличие от митоза, является важным элементом полового размножения . При мейозе образуются клетки, содержащие лишь один набор хромосом, что делает возможным последующее слияние половых клеток (гамет) двух родителей. По сути, мейоз является разновидностью митоза. Он включает два последовательных деления клетки, однако хромосомы удваиваются только в первом из этих делений. Биологическая сущность мейоза заключается в уменьшении числа хромосом в два раза и образовании гаплоидных гамет (то есть гамет, имеющих по одному набору хромосом).
В результате мейотического деления у животных образуются четыре гаметы . Если мужские половые клетки имеют примерно одинаковые размеры, то при образовании яйцеклеток распределение цитоплазмы происходит очень неравномерно: одна клетка остаётся крупной, а три остальных настолько малы, что их почти целиком занимает ядро. Эти мелкие клетки служат лишь для размещения избыточного генетического материала.
Мужские и женские гаметы сливаются, образуя зиготу . Хромосомные наборы при этом объединяются (этот процесс называется сингамией ), в результате чего в зиготе восстанавливается удвоенный набор хромосом – по одному от каждого из родителей. Случайное расхождение хромосом и обмен генетическим материалом между гомологичными хромосомами приводят к возникновению новых комбинаций генов, повышая генетическое разнообразие. Образовавшаяся зигота развивается в самостоятельный организм.
В последнее время проводились эксперименты по искусственному слиянию клеток одного или разных видов. Наружные поверхности клеток склеивались вместе, а мембрана между ними разрушалась. Таким образом удалось получить гибридные клетки мыши и цыплёнка, человека и мыши. Однако при последующих делениях клетки теряли большинство хромосом одного из видов.
В других экспериментах клетка разделялась на компоненты, например, ядро, цитоплазму и мембрану. После этого компоненты различных клеток снова соединяли вместе, и в результате получалась живая клетка, состоящая из компонентов клеток разных видов. В принципе, опыты по сборке искусственных клеток могут оказаться первым шагом на пути к созданию новых форм жизни.
Это расщепление, разделяющее кинетохоры, не зависит от других событий митоза и происходит даже в хромосомах, не прикрепленных к митотическому веретену. Оно позволяет полярным силам веретена, действующим на метафазную пластинку, начать перемещение каждой хроматиды к соответствующим полюсам веретена со скоростью порядка 1 мкм/мин. Если бы не было нитей веретена, то хромосомы расталкивались бы во все стороны, но благодаря наличию этих нитей один полный набор дочерних хромосом собирается у одного полюса, а другой -- у другого. Во время движения к полюсам хромосомы обычно принимают V-образную форму, причем вершина их обращена к полюсу. Центромера располагается у вершины, и, сила, заставляющая хромосому двигаться к полюсу, приложена к центромере. Хромосомы, утратившие центромеру во время митоза совсем не движутся
5) Телофаза начинается после того, как дочерние хромосомы, состоящие из одной хроматиды, достигли полюсов клетки. На этой стадии хромосомы вновь деспирализуются и приобретают такой же вид, какой они имели до начала деления клетки в интерфазе (длинные тонкие нити). Вокруг них возникает ядерная оболочка, а в ядре формируется ядрышко, в котором синтезируются рибосомы. В процессе деления цитоплазмы все органоиды распределяются между дочерними клетками более или менее равномерно. На этом завершается деление ядра, называемое также кариокинезом; затем происходит деление тела клетки, или цитокинез.
Таблица 2. Фазы митоза
В большинстве случаев весь процесс митоза занимает от 1 до 2 ч. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму; она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки. Материал клеточной пластинки вырабатывается эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточной пластинки цитоплазматическую мембрану, и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки.
Частота митозов в разных тканях и у разных видов резко различна. Например, в красном костном мозге человека, где в каждую секунду образуется 10 000 000 эритроцитов, в каждую секунду должно происходить 10 000 000 митозов.

Один из древнейших храмов Московского Кремля стоит на краю Соборной площади на бровке Боровицкого холма. Много веков...

10 августа в возрасте 63 лет умер Александр Коровников - депутат Госдумы от Новгородской области, член фракции "Единая...

Как вкусно и необычно приготовить груши на зиму, чтобы угощать всю семью морозными днями ароматным и витаминным летним...

[арам. ; греч. Βαρνάβας] († ок. 57), ап. от 70 (пам. 11 июня и в Соборе 70 апостолов). Считается основателем Кипрской...

Ну что, пора начинать готовиться к новогодним праздникам? Даже не знаю, существует ли более рождественское основное...

Увиденная во сне невеста символизирует наивность и чистоту помыслов в реальной жизни. Она может быть в обуви, фате, на...

Я хочу подарить Вам самую сильную молитву от колдовства и порчи, что есть у меня. Я никогда и нигде не видела...

несколько толкований. церковь – Вас икона, значит, ваши к веселью и жизненному опыту. (Например, и...

Говоря о покупке недвижимости за границей, часто упоминают о достопримечательностях страны, о ее истории и...

Копченый сыр прекрасно сочетается с сырой морковью, и именно с таким сыром салат получается более нежным....

Профессионалы кулинарии небезосновательно считают, что луковый соус – это универсальная подлива к мясным,...

Калорийность: 1450 Белки/100г: 11 Углеводы/100г: 15 Если вы любите чизкейк, но содержание калорий в этом...

Выписка из Единого государственного реестра юридических лиц (далее – ЕГРЮЛ) является документом, содержащим...

Это очень успешный человек, подающий надежды.К вершинамВячеслав Лысаков - самый обычный, из народа, с добрым...

10 августа в возрасте 63 лет умер Александр Коровников - депутат Госдумы от Новгородской области, член фракции...

Как вкусно и необычно приготовить груши на зиму, чтобы угощать всю семью морозными днями ароматным и витаминным...